РИБОСОМА, крупный
внутриклеточный макромолекулярный ансамбль, ответственный за синтез полипептидной
цепи из аминокислот (трансляцию); состоит из молекул РНК (т. наз. рибосомные
рибонуклеиновые кислоты, или рРНК) и белков.
Осн. масса рибосом локализована
в цитоплазме. В бактериальной клетке рибосомы составляют до 30% ее сухой массы: на
одну бактериальную
клетку приходится примерно 104 рибосом. В эукариотич. клетках (клетки всех
организмов, за исключением бактерий и синезеленых водорослей) относит. содержание
рибосом меньше, и их кол-во очень сильно варьирует в зависимости от белок-синтезирующей
активности соответствующей ткани или отдельной клетки.
В эукариотич. клетке все
рибосомы цитоплазмы (как мембрано-связанные, так и свободные) образуются в ядрышке;
считается, что там они неактивны. Эукариотич. клетка имеет также специальные
рибосомы в митохондриях (у животных и растений) и хлоропластах (у растений). Рибосомы этих
органелл отличаются от цитоплазматических размерами и нек-рыми функцион. св-вами.
Они образуются непосредственно в этих органеллах.
Различают два осн. типа
рибосом. Всем прокариотич. организмам (бактерии и синезеленые водоросли) свойственны
т. наз. 70S рибосомы, характеризующиеся коэф. (константой) седиментации ок. 70 единиц
Сведберга, или 70S (по коэф. седиментации различают и рибосомы др. типов, а также
субчастицы и биополимеры, входящие в состав рибосом). Их мол. м. составляет 2,5 ·
106, линейные размеры 20-25 нм. По хим. составу это рибонуклеопротеиды;
они состоят только из рРНК и белка (соотношение этих компонентов 2:1). Рибосомная
РНК в рибосомах присутствует гл. обр. в виде Mg-соли (по-видимому, частично и в виде
Са-соли); магния в рибосомах до 2% от сухой массы. Кроме того, в разл. кол-вах (до
2,5%) могут присутствовать также катионы аминов-спермина H2N(CH2)3NH(CH2)4NH(CH2)3NH2,
спермидина H2N(CH2)3NH(CH2)4NH2
и др.
Цитоплазма клеток всех
эукариотич. организмов содержит неск. более крупные 80S рибосомы. Их мол. м. ок. 4·106,
линейные размеры 25-30 нм, содержание белка в них значительно больше, чем в
прокариотической рибосоме (соотношение РНК: белок ок. 1:1). Рибосомная РНК 80S также
связана в осн. с Mg и Са и с небольшим кол-вом полиаминов (спермин, спермидин
и др.).
Хлоропласты и митохондрии
эукариотич. клеток содержат рибосомы, отличные от типа 80S. Рибосомы хлоропластов высших
растений принадлежат к истинному 70S типу. Митохондриальные рибосомы более разнообразны;
их строение находится в зависимости от таксономич. принадлежности организма
(т.е. от принадлежности к определенному виду, роду или семейству). Напр., митохондриальные
рибосомы млекопитающих существенно мельче типичных 70S рибосом; коэф. седиментации этих
рибосом составляет ок. 55S (т. наз. минирибосомы).
Рибосомыиз самых разнообразных
организмов (как прокариотич., так и эукариотич.) имеют сходное строение. Они
состоят из двух разделяемых субчастиц, или рибосомных субъединиц. При определенных
условиях (напр., при понижении концентрации Mg2 + в среде)
рибосома обратимо диссоциирует на две субчастицы с соотношением их мол. масс ок. 2:1.
Прокариотическая 70S рибосома диссоциирует на субъединицы с коэф. седиментации 50S
(мол. м. 1,5·106) и 30S (мол. м. 0,85·106). Эукариотическая
рибосома разделяется на субчастицы 60S и 40S. Две рибосомные субчастицы объединены
в полную рибосому строго определенным образом, предполагающим специфич. контакты их
поверхностей.
Как прокариотические, так
и эукариотические рибосомы содержат две разл. высокомол. рРНК (по одной на каждую
субчастицу) и одну относительно низкомол. рРНК в большой субчастице.
Рибосомные белки большинства
животных представлены в осн. умеренно основными полипептидами, хотя имеется
неск. нейтральных и кислых белков. Мол. м. рибосомных белков варьирует от 6
тыс. до 60 тыс. В прокариотической рибосоме малая субчастица (30S) содержит ок. 20,
большая (508)-ок. 30 разл. белков; в эукариотической рибосоме 40S субчастица включает
ок. 30 белков, а 60S-ок. 40 (обычно рибосомы не содержат двух или неск. одинаковых
белков). Рибосомные белки характеризуются глобулярной компактной конформацией
с развитой вторичной и третичной структурой; они занимают преим. периферич.
положение в ядре, состоящем из рРНК. В отличие от вирусных нуклеопротеидов в
структурно асим. рибосомном нуклеопротеиде рРНК не покрыта сплошной белковой
оболочкой, а в ряде мест образует пов-сть рибосом. Плотность упаковки рРНК в рибосомах достаточно
высока и приблизительно соответствует плотности кристаллич. упаковки гидратир.
полинуклеотидов.
По-видимому, рРНК определяет
осн. структурные и функцион. св-ва рибосом, в частности обеспечивает целостность рибосомных
субъединиц, обусловливает их форму и ряд структурных особенностей. Специфич.
пространств. структура рРНК детерминирует локализацию всех рибосомных белков,
играет ведущую роль в организации функцион. центров рибосом.
Рибосомный синтез белка-многоэтапный
процесс. Первая стадия (инициация) начинается с присоединения матричной РНК
(мРНК) к малой рибосомной субчастице, не связанной с большой субчастицей. Характерно,
что для начала процесса необходима именно диссоциированная рибосома. К образовавшемуся
т. наз. инициаторному комплексу присоединяется большая рибосомная субчастица.
В стадии инициации участвуют спец. инициирующий кодон (см. Генетический код),
инициаторная транспортная РНК (тРНК) и специфич. белки (т. наз. факторы
инициации). Пройдя стадию инициации, рибосома переходит к последоват. считыванию кодонов
мРНК по направлению от 5'- к 3'-концу, что сопровождается синтезом полипептидной
цепи белка, кодируемого этой мРНК (подробнее о механизме синтеза полипептидов
см. в ст. Трансляция). В этом процессе рибосома функционирует как циклически
работающая мол. машина. Рабочий цикл рибосомы при элонгации состоит из трех тактов:
1) кодонзависимого связывания аминоацил-тРНК (поставляет аминокислоты в рибосому),
2) транспептидации-переноса С-конца растущего пептида на аминоацил-тРНК, т.е.
удлинения строящейся белковой цепи на одно звено, 3) транслокации-перемещения
матрицы (мРНК) и пептидил-тРНК относительно рибосомы и переход рибосомы в исходное состояние,
когда она может воспринять след. аминоацил-тРНК. Когда рибосома достигнет специального
терминирующего кодона мРНК, синтез полипептида прекращается. При участии специфич.
белков (т. наз. факторов терминации) синтезир. полипептид освобождается из рибосомы.
После терминации рибосома может повторить весь цикл с др. цепью мРНК или др. кодирующей
последовательностью той же цепи.
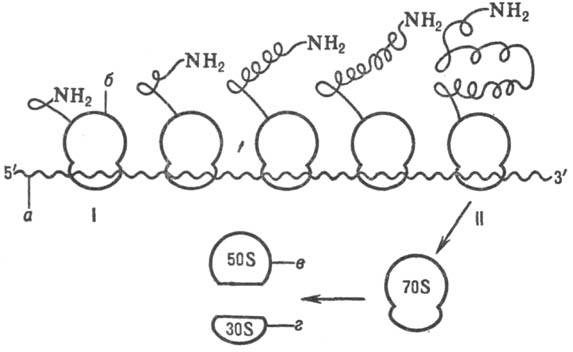
Схема синтеза полипептидной
цепи полирибосомой: I-начал о синтеза, II-окончание синтеза; а-мРНК,
б-рибосома, в-большая субъединица рибосомы, г-малая субъединица
рибосомы.
В клетках с интенсивной
секрецией белка и развитым эндоплазматич. ретикулумом значит. часть цитоплазматической
рибосомы прикреплена к его мембране на пов-сти, обращенной к цитоплазме. Эти рибосомы синтезируют
полипептиды, к-рые непосредственно транспортируются через мембрану для дальнейшей
секреции. Синтез полипептидов для внутриклеточных нужд происходит в осн. на
свободных (не связанных с мембраной) рибосомах цитоплазмы. При этом транслирующие рибосомы
не равномерно диспергированы в цитоплазме, а собраны в
группы. Такие агрегаты рибосом представляют собой структуры, где мРНК ассоциирована
со многими рибосомами, находящимися в процессе трансляции; эти структуры получили назв.
полирибосом или полисом.
При интенсивном синтезе
белка расстояние между рибосомами вдоль цепи мРНК в полирибосоме м. б. предельно коротким,
т.е. рибосомы находятся почти вплотную друг к другу. Рибосомы, входящие в полирибосомы,
работают независимо и каждая из них синтезирует полную полипептидную цепь (см.
рис.).
Лит.: Спирин А. С., Молекулярная биология. Структура рибосомы и биосинтез белка, М., 1986. П. Л. Иванов.