Рестриктазы
РЕСТРИКТАЗЫ (эндодезоксирибонуклеазы рестрикции), ферменты класса гидролаз, катализирующие гидролиз фос-фодиэфирных связей чужеродных ДНК в большинстве про-кариотических (бактерии и синезеленые водоросли) и нек-рых др. организмах и выполняющие тем самым "иммунную" ф-цию.
Распознавание чужеродной ДНК осуществляется в специ-фич. нуклеотидных последовательностях (сайтах), к-рые в собственной ДНК клетки "отмечены" благодаря модификации (чаще всего метилированию) спец. ферментами (напр., метилазами); иногда такой модифицирующей активностью обладают и сами рестриктазы.
По принятой номенклатуре рестриктазы обычно обозначают сокращенным назв. их продуцента, в к-ром первая буква-начальная буква назв. рода, две следующие-начальные буквы назв. вида и последующие - обозначение штамма и номера выделенной из него рестриктазы, напр.: HindIII—третья рестриктаза из Наето-philus influenzae d.
Рестриктазы подразделяют на 3 класса. К первому классу принадлежат ферменты (напр., ЕсоК из Escherichia coli К12), узнающие специфич. последовательность сайта, но разрывающие нить ДНК в произвольной точке (по-видимому, после образования комплекса с ДНК фермент неспецифически взаимод. с удаленной областью ДНК или передвигается вдоль нити ДНК). Ко второму классу принадлежат ферменты (напр., EcoRI), расщепляющие ДНК в строго определенной точке но отношению к сайту узнавания. К третьему классу относят ферменты промежут. типа (напр., EcoPI), разрывающие нить ДНК в неск. точках на разном удалении от сайта узнавания.
Наиб. значение имеют рестриктазы второго класса, широко применяемые в генетической инженерии и при установлении структуры ДНК. Эти ферменты (мол. масса, рН оптим. каталитич. активности и рI значительно варьируют в зависимости от источника) расщепляют обе нити ДНК. При этом разрыв осуществляет иногда одна молекула рестриктазы в обеих нитях, иногда-две молекулы фермента (каждая атакует лишь одну нить, как, напр., EcoRI). В большинстве случаев 3', 5'-фосфодиэфирные связи в ДНК расщепляются с образованием 5'-фосфатов на месте разрыва молекулы (см. Нуклеиновые кислоты), лишь немногие рестриктазы (напр., Nсil) образуют фрагменты с 3'-фосфатными группами. Все рестриктазы второго класса-Mg2+-зависимые ферменты. Для нек-рых рестриктаз (напр., EcoRI) установлена первичная структура.
Расщепление ДНК происходит обычно в пределах сайта узнавания, редко-на определенном расстоянии от него. При этом образуются фрагменты ДНК либо с ровными (тупыми) концами, либо с выступающими (липкими) 5'- или З'-концами (см. рис.). Рестриктазы с одинаковыми сайтами узнавания наз. изошизомерами.
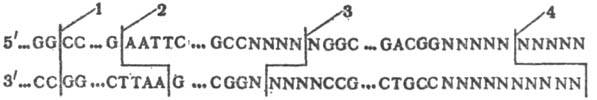
Схема расщепления ДНК разными рестриктазами: А, С, G и Т-соотв. дезок-сиаденозин, дезоксицитидин, дезоксигуанозин и дезокситимидин; N-любой дезоксинуклеозид, ферменты; 1-HalIII, 2-EcoI, 3-BglI, 4-HgaI.
Применение рестриктаз второго класса позволяет вырезать из ДНК определенные фрагменты, а также встраивать их в заданные места векторных молекул ДНК, т. е. направленно конструировать молекулы с новой генетич. информацией.
Лит.: Корнбсрг А., Синтез ДНК, пер. с англ., М., 1977, с. 298-305; Баев А. А., Таняшин В.И., Федор И., в сб.: Итоги науки и техники, сер. Молекулярная биология, т. 12, ч. 1. Генетическая инженерия, М., 1979; Kessler Оh, Neumaier P.S, Wolf W., "Gene", 1985, v. 33, № 1, p. 1-102.
А.И.Гуревич.