Клонирование ДНК
Развитие молекулярной генетики с 70-х гг. в значительной степени основано на разработке и совершенствовании методов анализа и манипулирования ДНК. Так называемая «генная (генетическая) инженерия» имеет практические приложения во многих областях. Например, были разработаны новые методы диагностики и лечения заболеваний и стало возможным проводить направленные изменения определенных свойств организма. Поскольку полностью исключить биологический риск невозможно, при использовании методов генной инженерии необходим осторожный подход. В этом и последующих разделах дан краткий обзор методов, используемых в генной инженерии.
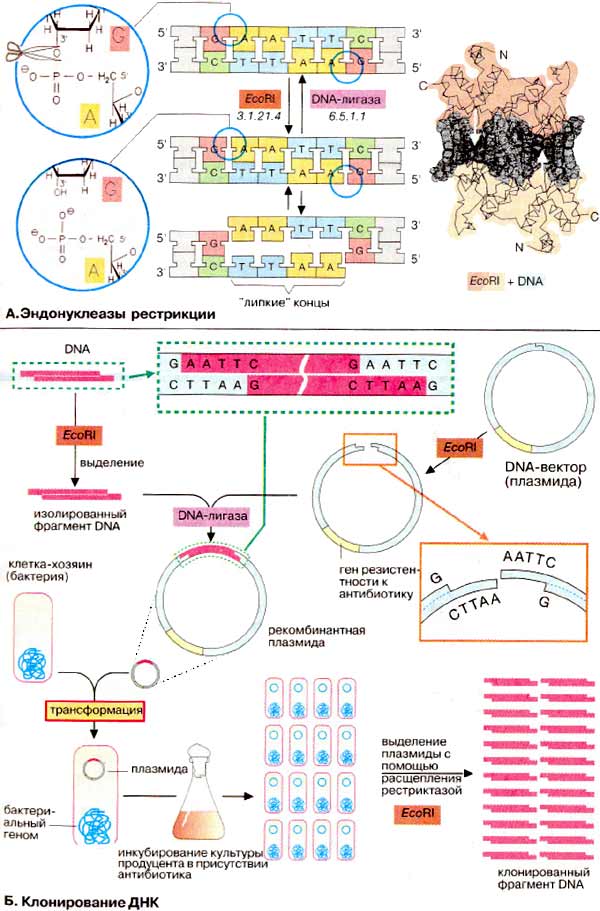
Большинство методик в генной инженерии включают выделение определенных фрагментов ДНК (DNA) и последующее их соединение с другими фрагментами для получения новых комбинаций генов. Для этих целей используются ферменты, которые специфически разрезают и вновь сшивают молекулы ДНК. Наиболее важной группой ферментов являются эндонуклеазы рестрикции (рестриктазы), катализирующие специфическое расщепление двунитевой ДНК. Известно большое число рестриктаз. Для их обозначения используются сокращенные названия микроорганизмов - продуцентов. В качестве примера рассмотрим EcoRI — эндонуклеазу, выделенную из Escherichia coli. Подобно многим другим рестриктазам, этот фермент расщепляет ДНК по палиндромной последовательности, т. е. короткому сегменту ДНК, в котором обе цепи при считывании в направлении 5'→3' имеют одинаковую последовательность. Для EcoRl это последовательность 5'-GAATTC-3'. Гомодимер EcoRI расщепляет фосфодиэфирные связи обеих цепей между G и А. Это приводит к образованию комплементарных «липких» концов (ААТТ), которые удерживаются вместе за счет спаривания оснований. Их, однако, можно легко отделить друг от друга путем небольшого нагревания. При охлаждении липкие концы гибридизуются вновь в правильной ориентации. Места расщепления можно соединить с помощью ДНК-лигазы.
Б. Клонирование ДНК
Обычно содержание в клетке какого-либо сегмента ДНК, например отдельного гена, очень незначительно. Поэтому для проведения экспериментов с фрагментами ДНК их необходимо многократно копировать (клонировать). В классической методике клонирования ДНК используется способность клеток бактерий поглощать и реплицировать короткие кольцевые молекулы ДНК, известные как плазмиды. Сначала клонируемый фрагмент ДНК вырезается из исходной ДНК с помощью рестриктазы. Для демонстрации метода на схеме показано расщепление с помощью EcoRI. На практике обычно используются два разных фермента. В качестве переносчика («вектора») служит плазмида с единственным участком, узнаваемым EcoRI. Кольцевая плазмида линеаризуется с помощью EcoRI и затем смешивается с изучаемым фрагментом ДНК. Поскольку фрагмент и вектор имеют одинаковые липкие концы, некоторые из молекул будут гибридизоваться таким образом, что клонируемый фрагмент окажется интегрированным в структуру вектора. Затем концы линейной молекулы ковалентно сшиваются с помощью ДНК-лигазы с образовании новой («рекомбинантной") плазмиды. При обработке большого количества клеток некоторые из них поглощают рекомбинантную плазмиду (так называемая трансформация). Трансформированные клетки реплицируют плазмиду вместе с собственным геномом. Обычно используют плазмиды, придающие трансформированной клетке устойчивость (резистентность) к определенному антибиотику. При инкубации популяции клеток в присутствии антибиотика будут реплицироваться только те клоны, которые содержат плазмиду. Из полученного клона выделяют плазмиду и после расщепления рестриктазой EcoRI получают множество копий клонированного фрагмента ДНК.