Биогенез матричных РНК
Процесс образования молекулы мРНК на матрице ДНК – биогенез мРНК – в прокариотических клетках представляется относительно простым и включает главным образом транскрипцию соответствующего гена при участии РНК-полимеразы. Во многих случаях первичным продуктом экспрессии гена является молекула мРНК, уже способная к функционированию, т.е. у прокариот транскрипция и трансляция являются сопряженными процессами. Биосинтез тРНК у прокариот из первичного тРНК транскрипта проходит стадию процессинга аналогично синтезу мРНК и тРНК у эука-риот (см. далее).
Биогенез мРНК у эукариот существенно отличается не только механизмом регуляции транскрипции, но и многоступенчатостью формирования активной молекулы. До открытия феномена сплайсинга (от англ. splicing – созревание, сращивание) мРНК было известно, что многие мРНК эукариот синтезируются в виде гигантских высокомолекулярных предшественников (пре-мРНК), которые уже в ядре подвергаются посттранскрипционному процессингу. Предполагали, что процессинг включает удаление длинных 5'- и 3'-концевых участков, которые якобы выполняют регуляторные функции. Как оказалось, ген эукариот является не непрерывной, а мозаичной структурой, содержащей наряду с кодирующими (экзоны) также некодирующие (интроны) последовательности. Фермент РНК-полимераза катализирует транскрипцию как экзонов (от англ. exit – выход, поскольку продукты транскрипции – участки мРНК – выходят из ядра в цитоплазму и выполняют функцию матрицы в синтезе белка), так и интронов с образованием гетерогенной ядерной РНК (гяРНК), называемой также первичным транскриптом. Термин «интроны» означает вставочные, нетранслирующие последовательности нуклеотидов в ДНК эукариот. Этот термин применим и к вставочным нуклеотидным последовательностям первичного РНК-транскрипта.
С открытием интрон-экзонного строения генов, характерного для эукариотических клеток, начался новый этап исследований на пути реализации генетической информации. Транскрипция гена, состоящего из чередующихся кодирующих и некодирующих нуклеотидных последовательностей, обеспечивала полное его копирование и приводила к синтезу РНК-предшественника. Поэтому было высказано предположение о существовании между транскрипцией и трансляцией еще одного важного звена – образования пригодной для трансляции «зрелой» молекулы мРНК. Этот этап получил название процессинга, или созревания, мРНК.
К настоящему времени считается установленным, что процессинг мРНК включает три основных процесса: 1) кэпирование – химическая модификация 5'-концевой последовательности мРНК; 2) сплайсинг – удаление некодирующих интронных последовательностей из мРНК и сшивание образующихся экзонов; 3) полиаденилирование – химическая модификация 3'-концевой последовательности мРНК (рис. 13.5).
В осуществлении каждого из указанных процесов специфическое участие принимает ряд белков и нуклеиновых кислот, хотя конкретные молекулярные механизмы этих превращений еще не полностью раскрыты. Все три указанных процесса имеют важное значение в формировании зрелой молекулы мРНК. Однако наибольший интерес исследователи проявляют к выяснению молекулярного механизма сплайсинга, который должен обеспечить, во-первых, постепенное и высокоточное вырезание интронов из первичного транскрипта и, во-вторых, сшивание образующихся фрагментов – экзонов – «конец в конец». Любые отклонения или смещения границ в процессе вырезания интронов и сшивания экзонов даже на один нуклеотид могут привести не только к глубокому искажению смысла в кодирующих последовательностях, но и к нарушению передачи генетической информации и развитию патологии.
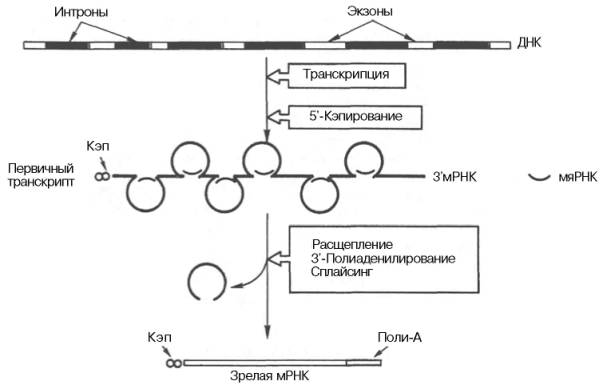
Рис. 13.5. Биогенез мРНК у эукариот.
Последовательность нуклеотидов в молекуле мРНК обычно начинается с пары 5'-ГУ и заканчивается парой АГ-3'. Эти последовательности, вероятнее всего, служат сайтами (местами) узнавания для ферментов сплайсинга. Поскольку 5'-ГУ ... АГ-3' последовательности не открыты в молекулах предшественников тРНК, было высказано предположение о существовании по меньшей мере двух типов ферментов сплайсинга; одного для мРНК и другого для тРНК. Имеются, кроме того, достоверные данные о том, что интроны часто оказываются длиннее экзонов и что внутри гена на интроны приходится значительно большая часть нуклеотид-ных пар. Подсчитано, например, что ген овальбумина содержит 7 интронов, в общей сложности насчитывающих 7700 пар оснований, в то время как сформировавшаяся после сплайсинга мРНК насчитывает всего 1859 оснований. Почти во всех эукариотических клетках синтезированные на структурных генах первичные транскрипты подвергаются процессингу, прежде чем выполнят свои уникальные функции в белковом синтезе. Во многих случаях процессинг имеет место главным образом в ядре, хотя этот процесс продолжается и после транспортировки молекул РНК из ядра в цитоплазму: например, терминальные реакции полиаденилирования и метилирования остатков нуклеозидов.
Химический смысл кэпирования сводится к присоединению остатка 7-метилгуанозина посредством трифосфатной группы к 5'-концу молекулы транскрипта, метилированию 2'-ОН-группы первого и второго нуклеотидов на 5'-конце мРНК. Полиаденилирование 3'-конца первичного транскрипта включает ряд стадий и участие эндонуклеазы и полиаденилатполимеразы. Эндонуклеаза расщепляет мРНК вблизи специфической сигнальной последовательности (5')ААУААА(3'), отличающейся высокой консервативностью. Полиаденилатполимераза синтезирует поли-А-конец (от 20 до 250 нуклеотидов) начиная с точки распада.
Функции 5'-кэп и 3'-поли-А раскрыты недостаточно полно. Показано, что 5'-кэп, соединяясь со специфическим белком, принимает участие в связывании мРНК с рибосомой, способствуя инициации синтеза белка. Допускают, что основное назначение 5'-кэп и поли-А – защита мРНК от энзиматического распада. Известно также, что не все цитоплазматические мРНК содержат участки поли-А на 3'-концах и что в цитоплазме клеток животных происходит как присоединение, так и удаление участка поли-А из молекулы мРНК. Следует отметить, что размер молекулы цитоплазма-тической мРНК даже после удаления 3'-поли-А оказывается все же намного большим, чем требуется для синтеза кодируемого белка. В частности, размер мРНК белка глобина (эритроциты кролика) составляет 550 нуклео-тидов, в то же время кодирующий участок состоит из 430 нуклеотидов (размер поли-А – 40 нуклеотидов). Другой пример: размер мРНК тяжелого иммуноглобулина (из клеток миеломы мышей) составляет 1800 нуклеотид-ных остатков, а кодирующая часть – 1350 нуклеотидов (размер поли-А – 150–200 нуклеотидов). Интересно, что большинство указанных процессов, если не все, могут регулироваться независимо, изменяя уровень экспрессии гена. Более того, даже после завершения формирования мРНК изменения ее стабильности могут оказывать существенное влияние на экспрессию гена.
В последние годы интенсивно исследуются структура и назначение нетранслируемых участков генов – интронов. Они различаются по числу, размерам и топографии. Показано, например, что ген сывороточного альбумина хотя и содержит всего 6 интронов, но на их долю приходится до 80% этого гена; интроны имеют размеры от 90 до 20000 нуклеотидных пар. Ген коллагена содержит более 50 интронов. Исключение составляют лишь гены, кодирующие гистоны, не содержащие интронных структур. Различают 4 класса интронов. Первый класс открыт как в ядерных, так и в митохондриальных генах, кодирующих рибосомные рРНК; второй класс интронов открыт в первичных транскриптах митохондриальных матричных мРНК. Оказалось, что оба эти класса интронов не нуждаются ни в источнике энергии, ни в участии ферментов, но наделены способностью самосплайсинга. Третий – самый большой класс интронов обнаружен в первичных транскриптах ядерных мРНК, подвергающихся созреванию. Сплайсинг требует наличия комплекса белков и особой группы клеточных РНК, названных малыми ядерными РНК (мяРНК). Выделено и охарактеризовано 5 групп богатых уридином мяРНК, соответственно обозначаемых U1, U2, U4, U5 и U6, размерами от 100 до 200 нуклеотидов. Комплексы мяРНК и белков, названные малыми ядерными нуклеопро-теинами, объединяются в единую систему – сплайсосому, координирующую весь процесс сплайсинга. Предполагают, что мяРНК соединяются с обеими концами интрона, способствуя формированию специфической конформации, необходимой для узнавания ее участвующими в процессе ферментами, сближению двух экзонов, удалению интронов и воссоединению кодирующих экзонов. Четвертый класс интронов открыт в ряде тРНК. Сплайсинг этой группы интронов требует доставки энергии и присутствия эндонуклеаз и лигаз, катализирующих соответственно разрыв фосфодиэфирных связей с 5'- и 3'-концов интрона и соединяющих два экзона.
Укажем также на весьма интересные и новые данные о существовании в структуре мРНК-предшественника, помимо экзонов и интронов, особых, так называемых альтернативно сплайсируемых, последовательностей. Выявлены примеры неоднозначного протекания сплайсинга для ряда генов. Результат альтернативного сплайсинга – появление нескольких продуктов при экспрессии одного гена. Так, получены доказательства, что экспрессия одного и того же гена тропомиозина позволяет получить семь изоформных белков, специфичных для разных групп мышц (гладких и поперечнополосатых) или для фибробластов и миобластов. В то же время известны примеры формирования одного белкового продукта (например, олиго-мерного фермента глюкозо-6-фосфатдегидрогеназы) при экспрессии двух разных генов. Все эти данные свидетельствуют о том, что альтернативный сплайсинг может играть существенную роль в функционировании генома клеток высших организмов.
В нетранскрибируемых последовательностях генома перед экзон-интронами открыты специфические участки, названные промоторами, а также энхансерами (повышающие уровень транскрипции) и силан-серами (ослабляющие уровень транскрипции). При взаимодействии с белками они выполняют функции регуляторных сигналов при транскрипции. Этот способ регуляции широко используется клетками эукариот как в процессах дифференцировки, так и при индукции репрессии (см. главу 14).
Нельзя не упомянуть об открытии рибозимов, т.е. молекул РНК, выступающих в качестве катализатора. Пожалуй, это единственные из известных макромолекул, которые наделены как информационной, так и каталитической функцией. Открытие каталитических РНК поколебало само понятие «фермент». Оказалось, что некоторые РНК осуществляют посттранскрипционный процессинг, катализируя самосплайсинг, т.е. участвуют в разрезании и удалении интронов. Наделенные рядом свойств истинных и эффективных катализаторов рибозимы участвуют в двух типах реакций: в гидролизе (разрыве) фосфодиэфирной связи и в реакциях трансэтерификации. В качестве субстрата могут служить, помимо собственного, предшественник (про-РНК) и другие молекулы РНК. Сейчас интенсивно изучается третичная структура рибозимов, а первичная и вторичная структуры ряда из них уже расшифрованы. Эти исследования, несомненно, интересные сами по себе, могут пролить свет и на пути развития биологической эволюции.
Для полного понимания молекулярных механизмов сложного процесса биогенеза мРНК предстоит решить множество вопросов. В частности, необходимо выделить в чистом виде и охарактеризовать белковые факторы, принимающие участие в этой регуляторной системе. Далее следует раскрыть механизмы узнавания промотора, терминации и антитерминации, избирательного метилирования, а также тонкие молекулярные механизмы регуляции сплайсинга. Решение указанных проблем будет, несомненно, способствовать лучшему пониманию сущности механизмов регуляции экспрессии генов эукариотических клеток в норме и при патологии.