Матричные рибонуклеиновые кислоты
МАТРИЧНЫЕ РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ (информационные РНК, мРНК, иРНК), молекулы РНК, представляющие собой комплементарные (см. Комплементарность) копии участков значащих цепей генов (ДНК), в к-рых закодирована информация об аминокислотных последовательностях полипептидных цепей белков. Образуются в результате транскрипции (синтез РНК на ДНК-матрице с помощью фермента ДНК-зависимой РНК-полимеразы) и послед. процессинга первичного транскрипта - удаления избыточных РНК-последовательностей с обоих концов молекулы и некодирующих последовательностей РНК, а также сшивания кодирующих участков РНК друг с другом. мРНК переносят генетич. информацию от генов к рибосомам, где происходит трансляция (синтез полипептидных цепей на мРНК-матрице). При этом одну цепь мРНК могут одновременно транслировать неск. рибосом (т. наз. полирибосомы, или полисомы). мРНК всегда состоят из одной полинуклеотидной цепи, т. е. относятся к однотяжевым РНК. Как и любые др. РНК, мРНК включают остатки адениловой (А), гуаниловой (G), уридиловой (U) и цитидиловой (С) к-т, связанных между собой 3'-5'-фосфодиэфирными связями (ф-ла I). Вторичная структура мРНК представлена многочисл. двухспиральными участками ("шпильками"), образующимися в результате комплементарного спаривания оснований (А с U и G с С) одной и той же цепи; шпильки соединены между собой короткими однотяжевыми участками. При связывании с рибосомой макромолекулярная структура мРНК существенно изменяется, а двухспиральные ее участки в районе непосредств. контакта с рибосомой переходят в однотяжевое состояние.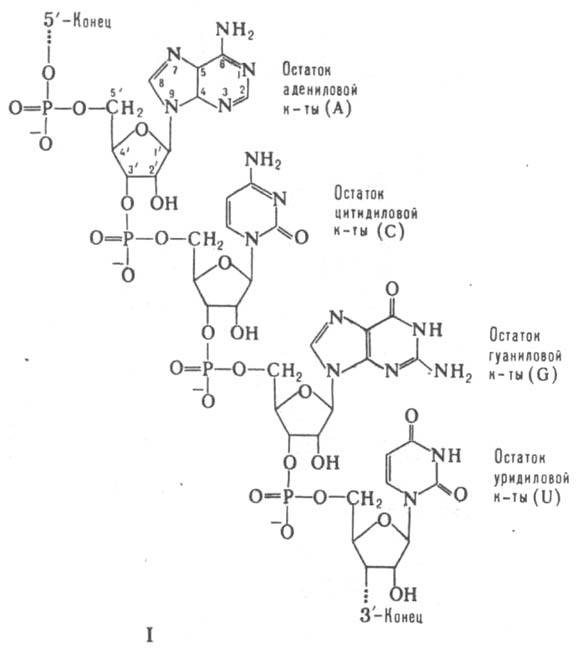
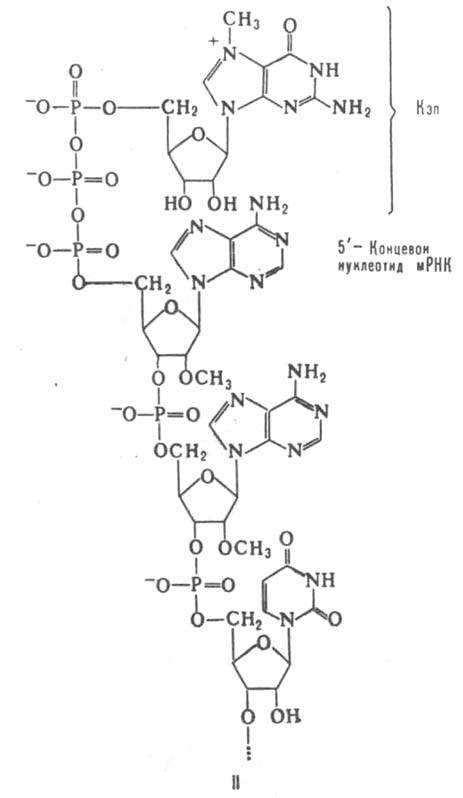
Полиаденилатные последовательности на 3'-концах мРНК часто используют также для получения их ДНК-копий с помощью олигомера тимидиловой к-ты в качестве затравки и фермента обратной транскриптазы. Эти копии в свою очередь используют для получения индивидуальных генов методами генетич. инженерии. Существование мРНК предсказано в 1956-58 А. С. Спириным и А. Н. Белозерским, а также независимо от них Э. Волкиным и Ф. Астрачаном. Экспериментально мРНК открыты в 1961 в лабораториях М. С. Мезелсона, Дж. Д. Уотсона и С. Спигелмена.
Литература
Информационные макромолекулы, пер. с англ., М., 1965; Спирин А. С., Молекулярная биология. Структура рибосомы и биосинтез белка, М., 1986; Льюин Б., Гены, пер. с англ., М., 1987. А. А. Богданов.
Синонимы/примеры:*
мРНК; матричная РНК; информационная РНК
*подобраны ИИ, возможны неточности
Ещё по теме
Транспортные РНК — строение и функции в биосинтезе белка
Транспортные РНК — строение и функции
Рибонуклеиновые кислоты — строение и функции
Молекулярная генетика — механизмы передачи генетической информации
Рибосомные рибонуклеиновые кислоты — строение и функции
Первичная структура нуклеиновых кислот
Биогенез матричных РНК — механизмы и процессы
Нуклеиновые кислоты — структура, функции, методы изучения
Механизмы биосинтеза РНК и экспрессии генов
РНК-полимеразы — строение и функции
Роль и типы РНК в генетике
Процесс транскрипции — от ДНК к РНК
Матричная РНК и синтез белка
Транскрипция — биосинтез РНК на матрице ДНК
Роль матричной РНК в синтезе белка
Биогенез рибосомных РНК — механизмы и регуляция
Вторичная структура нуклеиновых кислот
Химический состав нуклеиновых кислот — ДНК и РНК
Механизмы биосинтеза ДНК — роль ферментов и особенности репликации