Биосинтез РНК
Поток генетической информации называется экспрессией генов. Он включает процесс транскрипции – биосинтез матричных РНК (как и других типов клеточных РНК) на молекуле ДНК, и процесс трансляции – биосинтез белка на мРНК, т.е. генетическая информация ДНК реализуется путем программированного через мРНК синтеза белков, определяющих в конечном счете фенотипические признаки живых организмов. Подсчитано, что около 90–95% ДНК E. coli экспрессируется в мРНК, хотя большая часть последней не кодирует синтеза белка; небольшая часть ДНК кодирует синтез двух других клеточных РНК, т.е. рРНК и тРНК. Транскрипция, несмотря на кажущуюся схожесть с репликацией, в частности химическим механизмом, направлением синтеза и использованием матрицы, отличается рядом особенностей: не требует синтеза праймера, использует не всю молекулу ДНК, а только ее отдельные короткие сегменты (отдельные гены или группы генов) и, наконец, требует наличия только одной из цепей ДНК в качестве матрицы, которая полностью сохраняется (при репликации ДНК она сохраняется наполовину). Геном каждой клетки человека состоит из 3,5•109 пар оснований; они могут обеспечить кодирование более 1,5•106 пар генов. Однако имеющиеся данные о количестве и разнообразии белков в организме человека (около 100000) свидетельствуют о том, что значительная часть генома человека не транскрибируется и соответственно не переводится на аминокислотную последовательность белков. Известно также, что определенная часть нетранслируемого генома человека выполняет регуляторную функцию в процессе экспрессии генов. В молекуле ДНК различают, кроме уникальных неповторяющихся последовательностей, содержащих кодирующие гены, также множество повторяющихся последовательностей (повторы), биологический смысл которых до сих пор неясен (см. далее).
Современные представления о механизме синтеза РНК в клетках в значительной степени обязаны открытию в 1960 г. в двух лабораториях США (Дж. Хервиц и С. Вейс) особого фермента – РНК-полимеразы, катализирующей синтез РНК из свободных нуклеозидтрифосфатов. Фермент требует наличия ионов Mg2+или Мn2+ и одновременного присутствия всех 4 типов рибонуклеозидтрифосфатов (АТФ, ГТФ, ЦТФ и УТФ). Самым удивительным свойством фермента оказалось то, что для включения нуклеотидов в РНК необходимо обязательное присутствие предобразо-ванной ДНК-матрицы . При тщательном изучении механизма синтеза РНК при участии РНК-полимеразы, называемой также ДНК-зависимой РНК-полимеразой (транскриптазой), было установлено, что молекула предобразованной ДНК, необходимая для реакции полимеризации, полностью определяет последовательность рибонуклеотидов во вновь синтезированной молекуле РНК. Другими словами, на матрице ДНК комплементарно строится полирибонуклеотид, являющийся копией первичной структуры ДНК, с той только разницей, что вместо тимидилового нуклео-тида ДНК в РНК включается уридиловый нуклеотид. Реакция синтеза РНК в общем виде может быть представлена следующим образом:
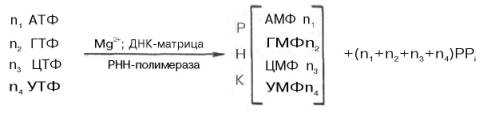
В синтезируемой молекуле РНК отдельные мононуклеотиды, как и в ДНК, связаны между собой 3'-5'-фосфодиэфирными мостиками. Кроме того, сам механизм действия фермента РНК-полимеразы во многом совпадает с таковыми ДНК-полимеразы: синтез также идет в направлении 5'–>3', цепь РНК имеет полярность, противоположную цепи предобразованной ДНК. Однако выявлены и существенные различия. РНК-полимераза Е. coli предпочтительнее функционирует в присутствии нативной двухцепочечной ДНК; в опытах in vitro обе цепи ДНК копируются РНК-полимеразой; in vivo транскрибируется, вероятнее всего, только одна цепь ДНК. Предполагают, что РНК-полимераза связывается с одной цепью нативной ДНК в определенной точке, вызывая расплетение биспиральной структуры на ограниченном участке, где и происходит синтез РНК. Данные свидетельствуют, что у E. coli, скорее всего, имеется единственная ДНК-зависимая РНК-полимераза, которая катализирует синтез всех типов клеточных РНК.
РНК-полимераза Е. coli изучена наиболее подробно. Это олигомерный фермент, состоящий из двух одинаковых α-субъединиц (мол. масса 36000), двух разных β (β1и β2)-субъединиц (мол. масса соответственно 151000 и 155000), ω-субъединицы (мол. масса 11000) и σ-субъединицы; общая мол. масса фермента около 390000. Считают, что функция σ-субъединицы (σ-фактор) – узнавание определенного участка на матрице ДНК, названного промотором, к которому присоединяется РНК-полимераза. В результате образуется так называемый открытый комплекс фермента с ДНК: двух-цепочечная структура ДНК раскрывается («плавится»). Далее на одной из нитей ДНК, как на матрице, синтезируется мРНК; синтез заканчивается в определенной точке в конце гена или прерывается под действием особых белков. Другим субъединицам фермента приписывают функцию инициации биосинтеза РНК (α-субъединицам) и основную каталитическую функцию (связывание субстратов и элонгация синтеза) – β-субъединицам. Кроме того, открыт ряд белков, принимающих участие в механизме синтеза РНК в клетке. В частности, исследуется природа репрессорных белков и белка-терминатора (ρ-фактора). Последний обладает способностью обратимо связываться с терминирующими участками ДНК (так называемые стоп-сигналы транскрипции), выключая действие РНК-полимеразы. При отсутствии этого белка образуются исключительно длинные цепи РНК.
У эукариот открыты три разные РНК-полимеразы (I, II и III) с большой молекулярной массой (от 500000 до 600000), каждая из которых наделена специфической функцией. РНК-полимераза I ответственна за синтез только рибосомных РНК (рРНК), точнее одного-единственного прерибосомного РНК-транскрипта, предшественника 5,8S, 18S и 28S рРНК; фермент связывается с разными промоторными участками. РНК-полимераза II – основной фермент, катализирующий синтез матричной РНК (мРНК). Он наделен способностью распознавать огромное множество промоторных участков, многие из которых имеют специфические ключевые последовательности, являющиеся местами (сайтами) связывания транскрипционных белковых факторов. РНК-полимераза III катализирует преимущественно синтез транспортных РНК (тРНК), а также 5S рРНК и ряда других низкомолекулярных РНК со специфической функцией. У эукариот работу РНК-полимеразы обеспечивает множество регуляторных белков (факторы транскрипции), объединенных вместе с ферментом в единый транскрипционный комплекс. В частности, открыты транскрипционные факторы типа J, активные только в виде идентичных димеров (J1J1 или J2J2) или разных димеров (J1J2); эти факторы кодируются отдельными генами и сами запускают работу ряда генов, регулирующих клеточное деление. В результате мутации генов, кодирующих синтез транскрипционных факторов, резко повышается прочность связывания J-факторов с ДНК, что обычно приводит к нерегулируемому опухолевому росту клеток.