Рекомбинация генетическая
РЕКОМБИНАЦИЯ ГЕНЕТИЧЕСКАЯ, реорганизация ге-нетич. материала, обусловленная обменом отдельными сегментами (участками) двойных спиралей ДНК.
Рекомбинация генетическая-главный фактор непостоянства генома, основа большинства его изменений, обусловливающая естеств. отбор, микро- и макроэволюции.
Различают два осн. типа рекомбинации генетической: 1) "законную" (общую, или гомологичную), при к-рой происходит обмен гомологичными (одинаковыми) участками молекул ДНК; 2) "незаконную" (негомологичную), в основе к-рой лежит обмен негомологичными участками ДНК.
Если обмен между разными молекулами ДНК осуществляется только в участках со строго определенными нуклеотидными последовательностями, рекомбинацию генетическую наз. сайт-специфичной, если в любых местах молекулы ДНК-сайт-кеспецифичной.
Законная рекомбинация генетическая обычно сайт-неспецифична, хотя довольно часто у бактерий и высших организмов она может проявлять черты сайт-специфичности, т. е. избирательности к определенным нуклеотидным последовательностям ДНК (т. наз. горячие точки рекомбинации). Такие последовательности резко повышают частоту рекомбинации генетической в тех участках генома, в к-рых они локализованы. Незаконная рекомбинация генетическая может быть как сайт-неспецифичной, так и весьма специфичной относительно участка обмена.
Законная рекомбинация генетическая наблюдается, напр., между двумя копиями к.-л. хромосомы. У эукариот (все организмы, за исключением бактерий и синезеленых водорослей) наиб. типичен обмен участками гомологичных хромосом в мейозе (деление клеток, в результате к-рого происходит уменьшение числа хромосом в дочерних клетках-осн. стадия образования половых клеток). Этот обмен может происходить между плотно конъюгированными хромосомами на ранних стадиях развития яйца или сперматозоида. Реже-законная рекомбинация генетическая осуществляется при обычном делении клеток (с сохранением числа хромосом)-митозе.
У прокариот (бактерии и синезеленые водоросли), у к-рых отсутствует мейоз, а геном представлен только одной молекулой ДНК, законная рекомбинация генетическая сопряжена с такими естеств. формами обмена и переноса генетич. материала, как конъюгация (хромосомы из донорской клетки передаются в рециниентную через протоплазменный мостик-пиль), трансформация (ДНК проникает из среды через клеточную оболочку), трансдукция (передача ДНК осуществляется бактериофагом, или вирусом бактерий). У вирусов рекомбинация генетическая происходит при заражении ими клеток. После лизиса клетки обнаруживаются вирусы с рекомбинантными ДНК. У прокариот рекомбинацию генетическую осуществляют спец. клеточные белки (многие из них ферменты).
В основе мол. механизма законной рекомбинации генетической лежит принцип "разрыв-воссоединение" двух гомологичных молекул ДНК. Этот процесс (его наз. кроссинговер) включает неск. промежут. этапов: 1) узнавание участков; 2) разрыв и ре-ципрокное (крест-накрест) воссоединение молекул: замена одних цепей гомологичными; 3) устранение ошибок, возникающих в результате неправильного спаривания участков. Точка обмена может возникать на любом участке гомологичных нуклеотидных последовательностей хромосом, вовлекаемых в обмен. При этом в точке обмена обычно не происходит изменения нуклеотидных последовательностей. Точность разрыва и воссоединения чрезвычайно велика: ни один нуклеотид не утрачивается, не добавляется и не превращается в к.-н. другой.
Основой всех предложенных схем рекомбинации генетической послужила т. наз. модель Холлидея, согласно к-рой рекомбинация генетическая начинается с разрыва только одной из двух цепей спирали ДНК. Вслед за разрывом один конец цепи вытесняется другим концом, к-рый наращивается ДНК-полимеразой. Вытесненный конец разорванной цепи спаривается со второй молекулой ДНК (образуется т. наз. гетеродуплекс), в свою очередь вытесняя там участок одной из ее цепей. В конце концов одиночные гомологичные цепи обмениваются реципрокно. После этого первонач. этапа спаривания две гомологичные спирали ДНК удерживаются вместе благодаря перекрестному обмену цепями-по одной от каждой спирали (см. рис.). Точка перекрестка далее может мигрировать, в результате чего дополнительно образуются или растут гетеродуп-лексные участки на обеих молекулах ДНК.
Структура с перекрещенными цепями может существовать в разл. стереоизомерных формах, возникающих в результате вращения составляющих ее элементов относительно друг друга. Изомеризация, к-рая как и др. стадии рекомбинации генетической контролируется генетически, изменяет положение двух пар цепей: две ранее перекрещивавшиеся цепи становятся неперекрещивающимися и наоборот.
Для того чтобы вновь восстановились две отдельные спирали ДНК и тем самым прекратился процесс спаривания, в каждой из двух перекрещенных цепей должен произойти разрыв. Если он происходит до того, как прошла изомеризация, то две исходные спирали ДНК отделяются друг от друга так, что у каждой из них генетически перестроенной оказывается только одна цепь. Если же разрыв двух перекрещенных цепей происходит после изомеризации, то обе молекулы ДНК претерпевают полную реорганизацию: часть каждой исходной спирали оказывается присоединенной (ступенчатым соединением) к части другой спирали.
Законная генетическая рекомбинация приводит к возникновению новых комбинаций специфических аллелей (различной формы одного и того же гена, обусловливающие различные варианты развития одного и того же признака-группы
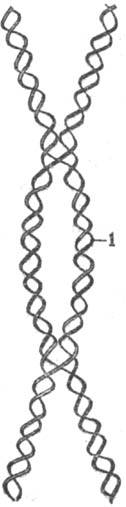
Схема спаривания двух гомологичных спиралей (одна из них обозначена жирной линией, другая—двойной); 1 — гетеродуплекс. крови, цвета глаз и др.), но не изменяет места расположения генов (локусов).
Незаконная рекомбинация генетическая имеет выраженный локальный характер. В этом случае весь процесс с его начальным этапом узнавания, к-рый сводит вместе две спирали ДНК, направляется особым рекомбинац. ферментом; спаривания оснований здесь не требуется (даже в тех случаях, когда это все-таки происходит, в процессе участвует не более неск. пар оснований). Интеграция транспозонов, плазмид и умеренных фагов в бактериальный геном может служить примером рекомбинации генетической этого типа. Подобный механизм существует также и в эукариотич. клетках (см. Мигрирующие генетические элементы).
При незаконной рекомбинации генетической в обмен вступают короткие специфич. нуклеотидные последовательности одной или обеих спиралей ДНК, участвующих в этом процессе. Таким образом такая рекомбинация генетическая изменяет распределение нуклеотидных последовательностей в геноме-соединяются участки ДНК, к-рые до этого не располагались в непрерывной последовательности рядом друг с другом. Подобный обмен гетерологич. участками ДНК приводит к возникновению вставок, делеций, дупликаций и транслокаций генетич. материала (см. Мутации).
У эукариот перемещения разных генетич. элементов, сопряженные с незаконной рекомбинацией генетической, осуществляются преим. не в мейозе, когда контактируют парные хромосомы, а во время обычных клеточных циклов (митозе). Незаконная рекомбинация генетическая играет важную роль в эволюц. изменчивости, т. к. благодаря ей осуществляются самые разнообразные, нередко кардинальные, перестройки генома и, следовательно, создаются предпосылки для качеств. изменений в эволюции данного организма.
Лит.: Зенгбуш П., Молекулярная и клеточная биология, пер. с нем., т. 1, М., 1982, с. 183-215; Страйер Л., Биохимия, пер. с англ., т. 3, М., 1985, с. 196-216. П.Л.Иванов.