Включение других углеводов в процесс гликолиза
Фруктоза. Установлено, что фруктоза, присутствующая в свободном виде во многих фруктах и образующаяся в тонкой кишке из сахарозы, всасываясь в тканях, может подвергаться фосфорилированию во фруктозо-6-фосфат при участии фермента гексокиназы и АТФ:
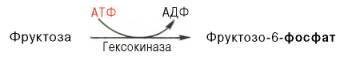
Эта реакция ингибируется глюкозой. Образовавшийся фруктозо-6-фос-фат либо превращается в глюкозу через стадии образования глюкозо-6-фосфата и последующего отщепления фосфорной кислоты (рис. 10.4), либо подвергается дальнейшим превращениям. Из фруктозо-6-фосфата под влиянием 6-фосфофруктокиназы и АТФ образуется фруктозо-1,6-бисфосфат:
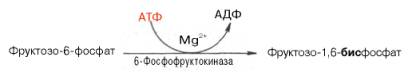
Далее фруктозо-1,6-бисфосфат может подвергаться дальнейшим превращением по пути гликолиза. Таков главный путь включения фруктозы в метаболизм мышечной ткани, почек, жировой ткани.
В печени, однако, для этого существует другой путь. В ней имеется фермент фруктокиназа, который катализирует фосфорилирование фруктозы не по 6-му, а по 1-му атому углерода:
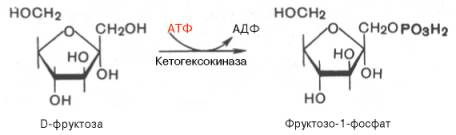
Эта реакция не блокируется глюкозой. Образовавшийся фруктозо-1-фосфат расщепляется затем под действием кетозо-1-фосфатальдолазы на диоксиацетонфосфат и D-глицеральдегид:
Фруктозо-1-фосфат <=> Диоксиацетонфосфат + D-глицеральдегид.
Образовавшийся D-глицеральдегид под влиянием соответствующей киназы (триокиназы) подвергается фосфорилированию до глицеральде-гид-3-фосфата. В этот же промежуточный продукт гликолиза переходит и дигидроксиацетонфосфат.
Существует врожденная аномалия обмена фруктозы, или эссенциальная фруктозурия, которая связана с врожденным недостатком фермента фрук-токиназы, т.е. в организме не образуется фруктозо-1-фосфат. В результате обмен фруктозы возможен только путем фосфорилирования до фрук-тозо-6-фосфата, но эта реакция тормозится глюкозой, вследствие чего фруктоза накапливается в крови. «Почечный порог» для фруктозы очень низок, поэтому фруктозурия обнаруживается уже при концентрации фруктозы в крови 0,73 ммоль/л.
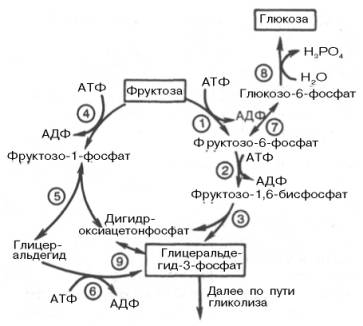
Рис. 10.4. Метаболизм фруктозы. 1 - гексокиназа; 2 - 6-фосфофруктоки-наза; 3 - фруктозобисфосфатальдолаза; 4 - кетогексокиназа; 5 - кетозо-1-фосфатальдолаза; 6 - триокиназа; 7 - глюкозофосфатизомераза; 8 - глюкозо-6-фос-фатаза; 9 - триозофосфатизомераза.
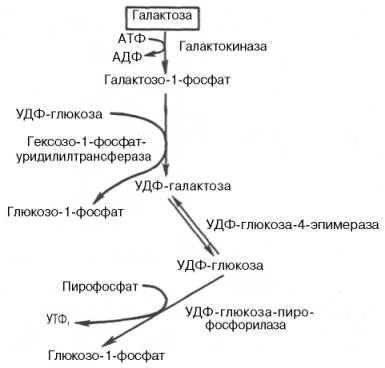
Рис. 10.5. Метаболизм галактозы.
Галактоза. Основным источником галактозы является лактоза пищи, которая в пищеварительном тракте расщепляется до галактозы и глюкозы (рис. 10.5).
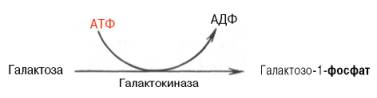
Обмен галактозы начинается с превращения ее в галактозо-1-фосфат. Эта реакция катализируется галактокиназой с участием АТФ:
В следующей реакции в присутствии УДФ-глюкозы фермент гексозо-1-фосфатуридилилтрансфераза катализирует превращение галактозо-1-фосфата в глюкозо-1-фосфат, одновременно образуется уридиндифосфат-галактоза (УДФ-галактоза):

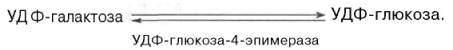
Образовавшийся глюкозо-1-фосфат в дальнейшем либо переходит в глюкозо-6-фосфат и далее подвергается уже известным превращениям, либо под влиянием фосфатазы образует свободную глюкозу, а УДФ-га-лактоза подвергается весьма своеобразной эпимеризации:
Затем УДФ-глюкоза-пирофосфорилаза катализирует расщепление УДФ-глюкозы с образованием глюкозо-1-фосфата:

О дальнейших превращениях глюкозо-1-фосфата см. ранее.
Одно из патологических состояний, возникающих в результате нарушения обмена углеводов,– это рецессивно наследуемое заболевание га-лактоземия. При этом заболевании общее содержание моносахаридов в крови повышается главным образом за счет уровня галактозы, достигая 11,1–16,6 ммоль/л. Концентрация глюкозы в крови существенно не изменяется. Кроме галактозы, в крови накапливается также галактозо-1-фос-фат. Галактоземия приводит к умственной отсталости и катаракте хрусталика. Возникновение данной болезни у новорожденных связано с недостатком фермента гексозо-1-фосфатуридилилтрансферазы. С возрастом наблюдается ослабление этого специфического нарушения обмена углеводов.