Контроль транскрипции
А. Функции регуляторных белков
Во всех клетках экспрессия генов контролируется регуляторными белками, которые связываются с определенным участком ДНК (DNA) и таким образом стимулируют пли подавляют транскрипцию гена (контроль транскрипции, см. с. 240). Действие регуляторных белков обратимо и, как правило, требует присутствия лиганда. Постоянно открывают все новые и новые регуляторные белки, в настоящее время известна, вероятно, только малая их часть. Несовершенна также их номенклатура. Как для белков, так и для участков ДНК, с которыми они связываются, используются различные наименования в зависимости от принципа действия. Регуляторный белок, который влияет на транскрипцию генов, называют фактором транскрипции. Белок, подавляющий транскрипцию, называют репрессором, а стимулирующий — индуктором. Последовательности ДНК, с которыми связываются регуляторные белки, называются регуляторными элементами. У прокариот регуляторные алименты, которые служат участками связывания РНК-полимеразы, называют промоторами, в то время как для репрессорных участков связывания употребляется название оператор. Регуляторные элементы, связывающие активирующие факторы, называют энхансерами (от англ. enhancer — усилитель), в то время как элементы, связывающие негативные (ингибирующие) факторы, — сайленсерами (от англ. silencer — успокоитель).
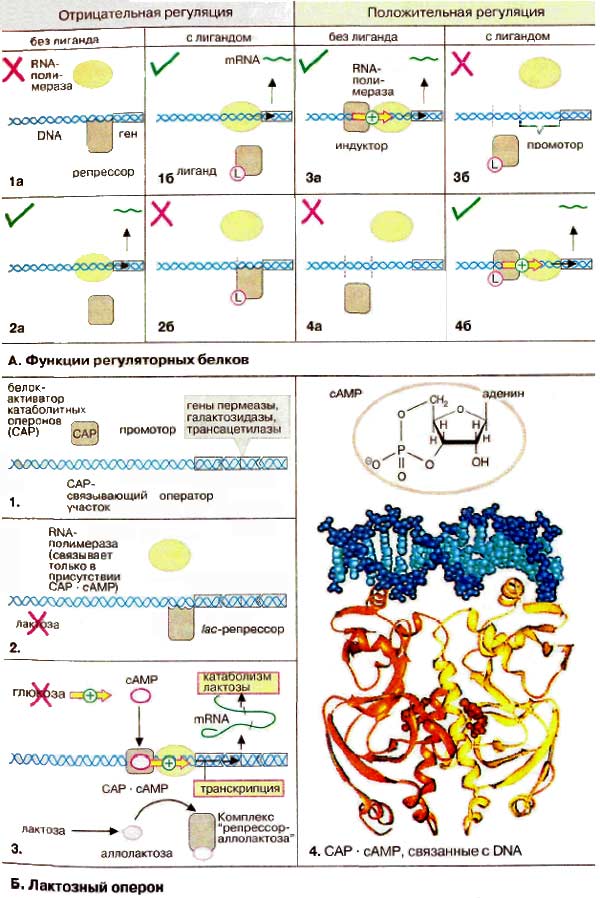
Многочисленные известные регуляторные белки можно разделить по механизму действия на четыре группы. Негативная генетическая регуляция, т. е. выключение соответствующих генов, может вызываться репрессорами. Некоторые репрессоры связываются с ДНК только в отсутствие специфического лиганда (1а). Комплекс репрессора с лигандом в этом случае теряет способность к связыванию и оставляет свободным участок промотора для присоединения РНК-полимеразы (1б). Часто свободный от лиганда репрессор не может связываться с ДНК, т. е. транскрипция подавляется только в присутствии лигандов (2а, 2б). Аналогично при позитивной генетической регуляции можно различать два случая. Если связывается только свободный индуктор, транскрипция подавляется соответствующими лигандами (3). Напротив, многие индукторы становятся активными только после образования комплекса с лигандом (4). К этой группе принадлежат, например, стероидные гормоны.
Б. Лактозный оперон
В качестве примера приведен лактозный оперон бактерии Е. coli (участок ДНК), который подвержен одновременно негативному и позитивному контролю. Оперон содержит структурные гены трех ферментов, которые необходимы для утилизации лактозы, и регуляторные элементы для управления транскрипцией оперона.
Так как лактоза превращается в клетке в глюкозу, экспрессия генов лактозного оперона не имеет смысла, когда глюкоза присутствует в клетке. Действительно гены транскрибируются только в отсутствие глюкозы и в присутствии лактозы (3). Регуляция достигается благодаря взаимодействию двух регуляторных белков. В отсутствие лактозы lac-penpeccop блокирует участок промотора (2). При наличии лактозы она превращается в изомерную аллолактозу, которая связывается с белком-репрессором и тем самым вызывает диссоциацию репрессора и оператора (3). Тем не менее этого недостаточно для транскрипции структурных генов. Для связывания РНК-полимеразы необходим индуктор, белок-активатор катаболитных оперонов (САР от англ. catabolite activator protein), который связывается с ДНК только в комплексе с цАМФ (cAMP). Сигнал голодания возникает только в отсутствие глюкозы.
Взаимодействие комплекса САР-цАМФ с ДНК представлено на рис. 4. Каждая субъединица димерного индуктора (желтого и оранжевого цвета соответственно) связывает молекулу цАМФ (красного цвета). Контакт с ДНК (голубого цвета) опосредуется двумя спиральными участками полипептидной цепи, специфически взаимодействующими с большой бороздкой на ДНК.