Молекулярные модели фотосистем
На схеме в упрощенной форме представлены бактериородопсин архебактерий Halobacterium halobium и фотосистема пурпурных бактерий Rhodopseudomonas viridis. Обе молекулы принадлежат к немногим трансмембранным белкам, структуры которых известны и могут служить в качестве модельных систем для подробного изучения фундаментальных механизмов энергетического обмена.
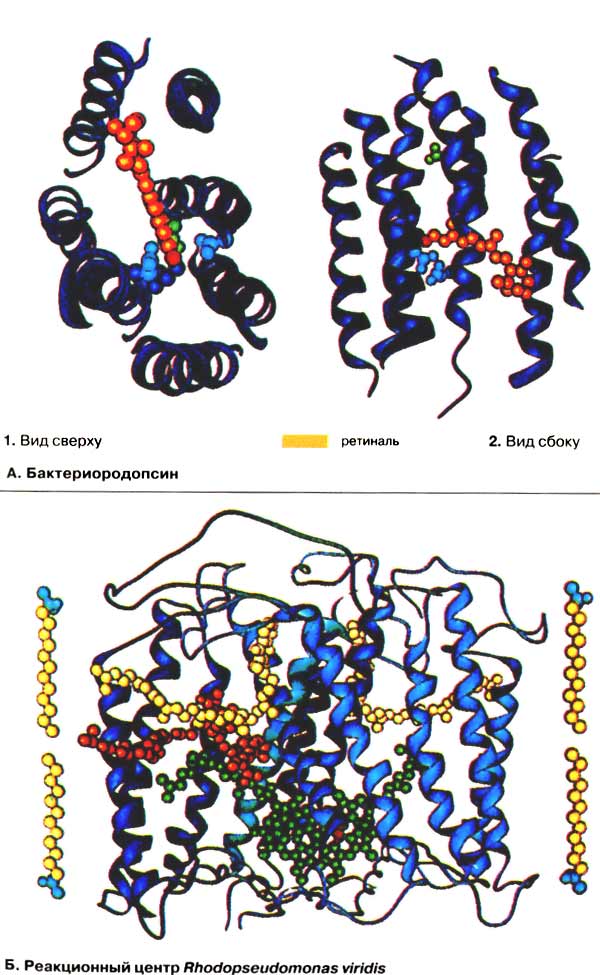
А. Бактериородопсин
Бактерии семейства Halobacterium растут при крайне высоких концентрациях соли, например в морской воде. Плазматическая мембрана этих бактерий содержит белок. подобный родопсину глаза и потому названный бактериородопсином. Этот белок способен непосредственно использовать энергию солнечного света для создания электрохимического градиента. В основе процесса, как и при зрительном процессе, лежит индуцируемая светом цис-транс-изомеризация ретиналя. Белковая часть молекулы бактериородопсина в основном состоит из 7 α-спиралей (голубого цвета), пронизывающих мембрану и образующих полый цилиндр. Остальная часть молекулы и боковые цепи аминокислот не представлены. Внутри цилиндра расположена молекула ретиналя (оранжевого цвета), ковалентно связанная альдегидной группой с ε-аминогруппой остатка лизина (красного цвета). В темноте ретиналь находится в полностью транс-форме, а альдиминная группа протонирована. При освещении ретиналь перегруппировывается в 13-цис-форму, а альдиминная группа отдает протон, который «откачивается» наружу двумя аспартатными остатками (светло-голубого цвета; на рис. 2 внизу). После возвращения ретиналя в полностью транс-форму альдиминная группа снова связывает протон. Внутриклеточный протон (2, вверху) переносится через другой аспартатный остаток (зеленого цвета) к ретиналю.
Б. Реакционный центр Rhodopseudomonas viridis
Фотосистема пурпурных бактерий Rhodopseudomonas viridis похожа по строению на фотосистему II высших растений. В отличие от растений в бактериальной системе донором электронов является не вода, а они поступают из электронпереносящей цепи, содержащей цитохром (на схеме не показано).
На схеме приведены только трансмембранные фрагменты бактериородопсина. О примерной толщине мембраны можно судить по липидным молекулам (слева и справа). Шесть трансмембранных спиралей из трех субъединиц (показаны окрашенными в различные тона голубого цвета) формируют внутримембранное пространство, которое наполнено цепями молекул пигментов (несколько пигментов, которые не принимают непосредственного участия в электронном транспорте, опущено). Принцип фотосинтетического электронного транспорта обсуждается на с. 130.
В Rh. viridis энергию света поглощают две соседние молекулы хлорофилла, образующие «специальную пару» (зеленого цвета, а ион Mg2+ — красного). Максимум поглощения этих молекул находится при 870 нм, поэтому бактериальный реакционный центр обозначается также P870 .
После возбуждения электрон переносится реакционным центром на смежную свободную от магния молекулу феофитина (оранжевого цвета) всего за несколько пикосекунд (1 пс = 10-12 с), а затем в течение примерно 200 пс передается на прочно связанный хинон QA (слева наверху, желтого цвета). В то же самое время снова заполняется электронная дыра в «специальной паре». Через примерно 0,2 мс возбужденный электрон достигает обмениваемого хинона QB (справа наверху, желтого цвета), с которым покидает фотосистему.