Механизм зрительного восприятия
В сетчатке глаза позвоночных содержатся два типа фоторецепторных клеток: палочки и колбочки. Палочки чувствительны к свету, а колбочки отвечают за восприятие цвета.
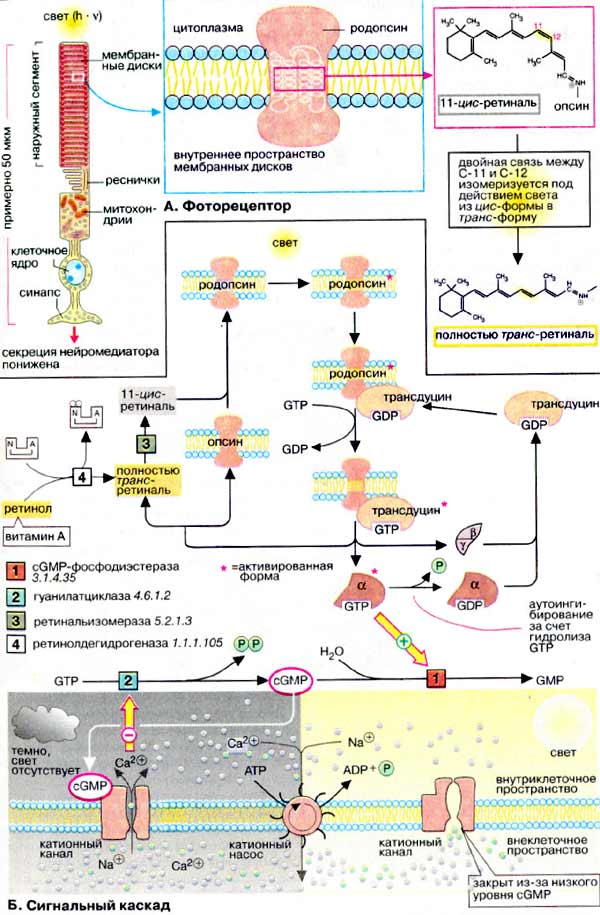
А. Фоторецептор
На рисунке схематически изображена одна из фоторецепторных клеток, палочка. Клетка состоит из двух основных частей, наружного и внутреннего сегментов. В дисках наружного сегмента (специализированных замкнутых мембранах) локализован родопсин, интегральный мембранный белок, включающий 7 трансмембранных тяжей. Такое строение характерно для большой группы сигналпереносящих рецепторных белков (рецепторов типа III, см. с. 372). Родопсин является светочувствительным хромопротеином. Помимо белковой части, опсина, молекула родопсина включает остаток 11-цис-ретиналя, связанный ковалентно с ε-аминогруппой остатка лизина. Родопсин обладает характерным спектром поглощения света с максимумом при 500 нм.
Поглощение молекулой родопсина кванта света индуцирует изомеризацию 11-цис-ретиналя в полностью транс-форму. В результате этой фотохимической реакции изменяется геометрия ретиналя, а спустя 10 мс происходит аллостерический переход родопсина в его активную форму (родопсин*). Стимуляция родопсином* G-белка запускает каскад передачи сигнала, который побуждает зрительную клетку уменьшить выброс нейромедиатора (глутамата), вследствие чего биполярные нейроны, связанные со зрительными клетками, посылают измененный импульс, что воспринимается как зрительное возбуждение.
Б. Сигнальный каскад
G-белок палочек носит название трансдуцин. Связывание активированного светом родопсина* (метародопсина II) с ГДФ-трансдуцином катализирует обмен ГДФ (GDP) на ГТФ (GTP). Активная форма трансдуцина (ГТФ-трансдуцин) диссоциирует на комплекс β, γ-субъединиц и ГТФ-α*-субъединицу, которая активирует цГМФ-фосфодиэстеразу (сGΜΡ - фосфодиэстеразу) [1], связывая ингибиторную субъединицу фермента.
В отсутствие света концентрация цГМФ (cGMP) в колбочках поддерживается на сравнительно высоком уровне (70 мкМ). Этот вторичный мессенджер постоянно синтезируется гуанилатциклазой и гидролизуется цГМФ-фосфодиэстеразой. Активация фосфодиэстеразы (при освещении родопсина) вызывает быстрое (в течение нескольких мс) падение уровня цГМФ.
Спустя короткое время α-субьединица трансдуцина инактивируется за счет медленного гидролиза связанного ГТФ и ассоциирует с комплексом β, γ-субъединиц. Родопсин* распадается на опсин и полностью транс-ретиналь, который изомеризуется в цис-ретиналь под действием изомеразы [3]. После сборки родопсина молекула возвращается в исходное состояние.
В темноте (на схеме слева внизу) высокий уровень цГМФ в палочках поддерживается благодаря активности гуанилатциклазы. Поэтому цГМФ-зависимые катионные каналы плазматической мембраны остаются открытыми и катионы Na+ и Ca2+ беспрепятственно поступают в клетку. При этом зрительная клетка постоянно выбрасывает нейромедиатор глутамат в синаптическую щель.
При освещении (на схеме справа внизу) уровень цГМФ резко падает за счет активации фосфодиэстеразы*, что приводит к перекрыванию ионных каналов. Так как ионы Na+ и Ca2+ постоянно выкачиваются из клетки, концентрация их быстро падает. Это приводит к гиперполяризации клетки и останавливает выброс нейромедиатора. Снижение концентрации ионов Ca2+ инициирует активацию гуанилатциклазы, что влечет за собой быстрый подъем уровня цГМФ настолько, что ионные каналы открываются вновь.