Синапсы
А. Холинэргические синапсы
Передача сигналов между нейронами и от нейронов к мышечным клеткам (так называемая нейронейрональная и нейромышечная трансдукция) происходит в нервных окончаниях (синапсах). С помощью сигнальных веществ, медиаторов. Синапсы образованы мембранами двух контактирующих клеток, пресинаптической и постсинаптической которые разделены узкой синаптической щелью. Медиатор выделяется в синаптическую щель за счет зкзоцитоза, диффундирует к рецепторам постсинаптической мембраны, связывается с ними и передает сигнал соседней клетке. Белки-рецепторы — это лиганд-активируемые ионные каналы либо мембранные белки, которые управляют ионными каналами посредством G-белков (см. рис. 373).
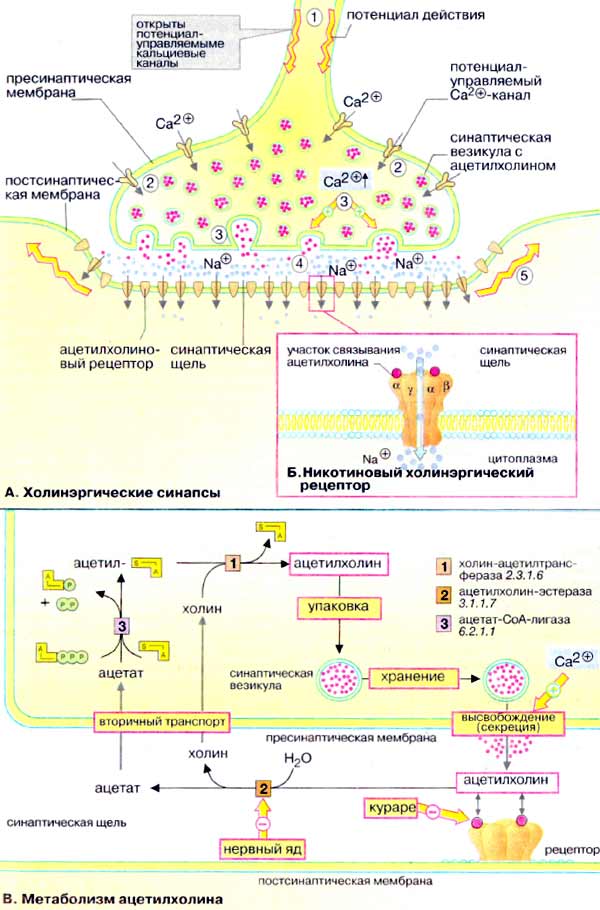
Ацетилхолин — нейромедиатор моторной концевой пластинки. Ацетилхолиновые рецепторы (никотиновый и мускариновый) — это лиганд-активируемые ионные каналы, которые открываются для прохождения ионов Na+ и К+. Никотиновые рецепторы (быстрые) локализованы главным образом в месте контакта аксонов со скелетными мышцами. Мускариновые рецепторы (медленные) локализованы в головном мозге, секреторных клетках, гладких и сердечных мышцах.
Процесс передачи сигнала включает следующие этапы. Потенциал действия достигает пресинаптической мембраны (1). Это вызывает открывание потенциал-управляемых Сa2+-каналов (2). Ионы Са2+ проникают из внеклеточного пространства в клетку, их уровень в синапсе резко увеличивается, что инициирует процесс экзоцитоза. Синаптические везикулы выделяют содержимое (ацетилхолин) в синаптическую щель (3). Молекулы ацетилхолина диффундируют через синаптическую щель, связываются с постсинаптическими рецепторами и активируют их (4). Поток ионов Na+ изменяет потенциал покоя постсинаптической мембраны нервной или мышечной клетки настолько, что открываются соседние потенциал-управляемые Na+ каналы и возникает потенциал действия (5, см. рис. 341).
Б. Никотиновый холинэргический рецептор
Наиболее детально изучен рецептор ацетилхолина, активируемый никотином. Это трансмембранный комплекс из пяти субъединиц (α2βδγ, 250-270 кДа), образующий лиганд-активируемый (хемовозбудимый) ионный канал, проницаемый для ионов Na+ и К+. Участки связывания ацетилхолина локализованы на внеклеточной части α-субъединиц. При связывании лиганда в центре молекулы формируется трансмембранный канал, входное отверстие которого имеет форму воронки диаметром около 2 нм. Предполагается, что в формировании канала принимают участие все пять субъединиц. Канал открывается на короткое время для прохождения ионов Na+ и К+. Считается, что открывание и закрывание канала происходит в результате аллостерических изменений в заряженных участках полипептидных цепей молекулы рецептора.
Рецептор может связывать различные лекарственные вещества: например, никотин действует как агонист ацетилхолина.
В. Метаболизм ацетилхолина
Ацетилхолин, уксуснокислый эфир холина, образуется в цитоплазме аксонов из ацетил-КоА и холина [1]. Нейромедиатор хранится в синаптических везикулах, в каждой везикуле содержится примерно 1000-10000 молекул ацетилхолина. После выделения из везикул ацетилхолин попадает в синаптическую щель. Избыток ацетилхолина расщепляется ацетилхолин-эстеразой [2]. Этот фермент имеет высокое число оборотов, что гарантирует быстрое удаление сигнального вещества. Продукты гидролиза, холин и уксусная кислота, активно захватываются пресинаптической частью синапса и используются для повторного синтеза ацетилхолина [3].
Соединения, блокирующие остаток серина в активном центре ацетилхолин-эстеразы [2], например токсин Е605, пролонгируют действие ацетилхолина и действуют как нейротоксины. Напротив, D-тубокурарин (яд кураре, которым индейцы пропитывали наконечники стрел) является конкурентным ингибитором ацетилхолина при связывании с рецептором.