Сохранение энергии на мембранах
Наряду с макроэргическими соединениями другим местом накопления химической энергии являются биологические мембраны. В технике система, работающая за счет разделения электрических зарядов непроводящим слоем, называется конденсатором. По принципу конденсатора функционируют биомембраны, разделяющие подобно изолирующему слою заряженные атомы и молекулы (ионы).
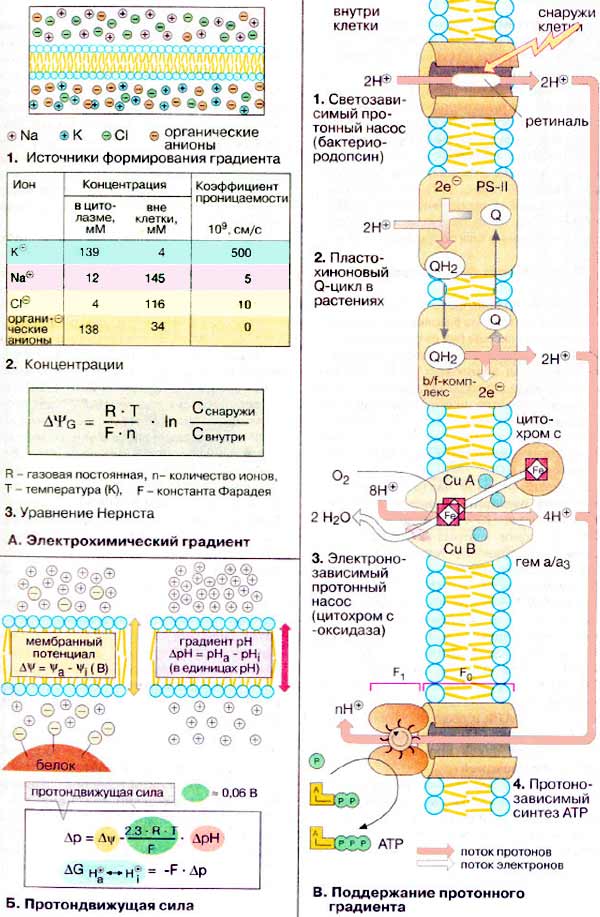
А. Электрохимический градиент
В то время как искусственная липидная мембрана для ионов практически не проницаема, биологические мембраны содержат «ионные каналы», по которым отдельные ионы избирательно проникают через мембрану. Проницаемость и полярность мембраны зависят от электрохимического градиента, т. е. от концентраций ионов по обе стороны мембраны (концентрационного градиента) и от разности электрических потенциалов между внутренней и внешней сторонами мембраны (мембранного потенциала).
В состоянии покоя клеток мембранный потенциал (потенциал покоя, см. с. 340) составляет от -0,05 до -0,09 В, т.е. на внутренней стороне плазматической мембраны преобладает избыток отрицательных зарядов. Потенциал покоя обеспечивается прежде всего катионами Na+ и K+, а также органическими анионами и ионом Cl- (1). Концентрации снаружи и внутри клетки и коэффициенты проницаемости этих ионов приведены в таблице (2).
Распределение ионов между внешней средой и внутренним объемом клетки описывается уравнением Нернста (3), где ΔΨG — трансмембранный потенциал (в вольтах, В), т.е. разность электрических потенциалов между двумя сторонами мембраны при отсутствии транспорта ионов через мембрану (потенциал равновесия). Для одновалентных ионов при 25оС множитель RT/Fn равен 0,026 В. Вместе с тем из таблицы (2) следует, что для ионов К+ ΔΨG примерно равно -0,09 В, т. е. величина того же порядка, что и потенциал покоя. Для ионов Na+, напротив, ΔΨG = +0,07 В, т.е. выше, чем потенциал покоя. Поэтому ионы Na+ поступают в клетку при открытии Na+-канала. Неравенство концентраций ионов Na+ и К+ постоянно поддерживается Na+/K+-АТФ-азой при расходовании АТФ.
Б. Протондвижущая сила
Ионы гидроксония («H+-ионы») также могут формировать электрохимический градиент. Такой протонный градиент имеет решающее значение для клеточного синтеза АТФ. Как и в случае других ионов, свободная энергия переноса протона (разность между электрохимическими потенциалами протонов на двух сторонах мембраны) зависит от градиента концентрации, т. е. от разности рН (ΔpH) по ту и другую стороны мембраны. Кроме того, определенный вклад вносит и трансмембранный потенциал ΔΨ. Обе эти величины формируют протондвижущую силу Δp, являющуюся мерой работы ΔΨG , которую может совершать H+-градиент. Таким образом, протонный градиент через внутреннюю митохондриальную мембрану дает примерно 24 кДж на моль переносимых ионов H+.
В. Поддержание протонного градиента
Протонные градиенты формируются различными способами. Необычным протонным насосом является бактериородопсин (1), использующий энергию света. При фотосинтезе восстановленный пластохинон (QH2) переносит протоны вместе с электронами через мембрану (Q-цикл, 2). Образование протонного градиента в дыхательной цепи также сопряжено с окислительно-восстановительным процессом. В комплексе III, по-видимому, как и при фотосинтезе, за перенос прогона ответствен Q-цикл (не показано). В цитохром с-оксидазе (комплекс IV, 3) H+-транспорт сопряжен с электронным потоком от цитохрома с на О2.
В каждом из этих случаев протонный градиент используется в синтезе АТФ АТФ-синтазой (4). АТФ-синтаза состоит из двух компонентов: канала протонов (F0) и управляемого им белкового комплекса (F1), который трансформирует энергию потока протонов через мембрану в химическую энергию АТФ.