Биосинтез антител
А. Вариабельность иммуноглобулинов
Несмотря на сходство своей основной структуры иммуноглобулины (Ig) чрезвычайно разнообразны. Считается, что в организме человека имеется примерно 108 различных вариантов антител. Вариабельность Ig относится как к легким, так и тяжелым цепям.
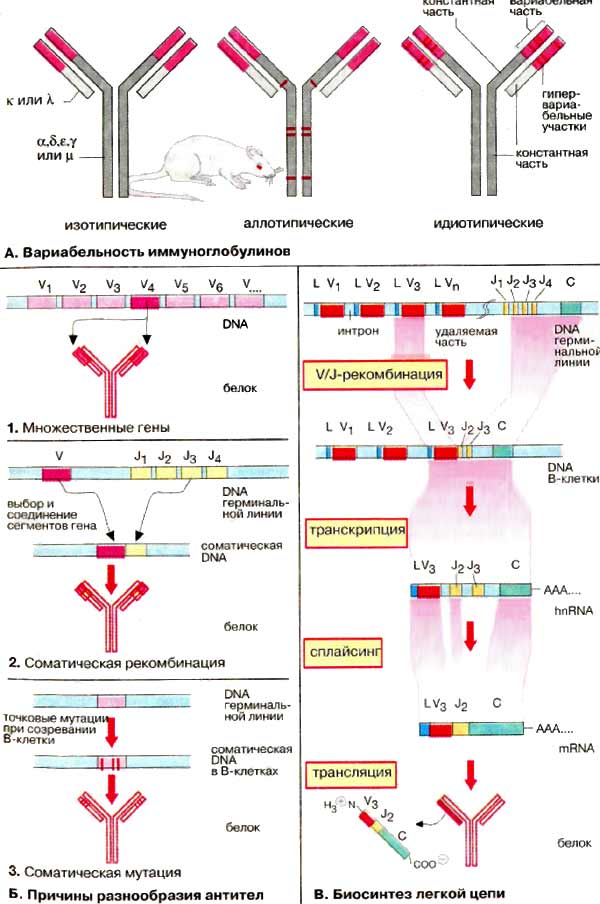
Имеется пять разновидностей тяжелых H-цепей (α, β, ε, γ и μ, см. с. 288), которые и определяют классы антител, и две разновидности легких L-цепей (κ и λ). Эти типы вариаций называют изотопическими. При биосинтезе Ig может происходить переключение плазматических клеток с продукции одного изотипа на другой («переключение генов»). Аллотипические вариации относятся к вариабельности аллелей в пределах вида, т. е. к генетически определяемым различиям одного индивидуума от другого. Идиотипические вариации определяют различия в антигенсвязывающем участке антител, Они касаются вариабельных доменов (на схеме розового цвета) легкой и тяжелой цепей. Некоторые их участки являются гипервариабельными (красного цвета на рисунке), т.е. их отличия особенно велики. Эти последовательности непосредственно участвуют в связывании антигена.
Б. Причины разнообразия антител
Исключительная вариабельность антител обусловлена тремя причинами.
1. Множественность генов. Имеется множество генов, кодирующих белки вариабельных доменов, однако выбирается и экспрессируется только один ген.
2. Соматические рекомбинации. Гены разделены на несколько сегментов, для которых имеются различные версии. Во время созревания В-клеток благодаря случайной комбинации сегментов возникают новые гены (мозаичные гены).
3. Соматические мутации. Во время дифференциации В-клеток и превращения в плазматические клетки происходят мутации в кодирующих генах. Таким образом, изначальные гены терминальной линии могут стать различными соматическими генами в индивидуальных клонах В-клеток.
В. Биосинтез легкой цепи
Рассмотрим основные особенности организации гена иммуноглобулина и его экспрессии на примере биосинтеза мышиной κ-цепи. Сегменты гена, кодирующие эту легкую цепь, обозначаются как L, V, J и С. Они локализованы на хромосоме 6 в ДНК (DNA) терминальной линии клеток мыши (у человека на хромосоме 2) и разделены друг от друга интронами различной длины.
Примерно 150 идентичных сегментов L гена кодируют сигнальный пептид (17-20 аминокислот) для секреции продукта. Наибольшая часть вариабельного домена (95 из 108 аминокислотных остатков) кодируется около 150 различными V-сегментами, расположенными рядом с L-сегментом. L- и V-сегменты всегда расположены парами, так называемым тандемом. Напротив, для J-сегмента (от англ. joining) существует максимально только пять вариантов. Они кодируют пептид из 13 аминокислотных остатков, который связывает вариабельную и константную части κ-цепи. Константная часть легкой цепи (84 аминокислоты) кодируется единственным С-сегментом.
Во время дифференциации В-лимфоцитов уникальная V/J-комбинация возникает в каждой В-клетке. Один из 150 сегментов L-V-тандема выбирается и связывается с одним из пяти J-сегментов. Это приводит к возникновению соматического гена, который значительно меньше по сравнению с геном терминальной линии. Транскрипция этого гена ведет к образованию гяРНК (hnRNA) для κ-цепи. Из этой РНК удаляются путем сплайсинга интроны и лишние J-сегменты. Зрелая мРНК (mRNA) содержит сегменты L-V-J-C и после транспорта в цитоплазму готова для трансляции. Последующие шаги биосинтеза Ig те же, что и для других мембранных или секреторных белков.