Молекулярная масса белков
Белки относятся к высокомолекулярным соединениям, в состав которых входят сотни и даже тысячи аминокислотных остатков, объединенных в макромолекулярную структуру. Молекулярная масса белков колеблется от 6000 (нижний предел) до 1000000 и выше в зависимости от количества отдельных полипептидных цепей в составе единой молекулярной структуры белка. Такие полипептидные цепи получили название субъединиц. Их мол. масса варьирует в широких пределах – от 6000 до 100000 и более.
Аминокислотный состав и последовательность аминокислот выяснены для многих тысяч белков. В связи с этим стало возможным вычисление их молекулярной массы химическим путем с высокой точностью. Однако для огромного количества встречающихся в природе белков химическое строение не выяснено, поэтому основными методами определения молекулярной массы все еще остаются физико-химические методы (гравиметрические, осмометрические, вискозиметрические, электрофоретические, оптические и др.). На практике наиболее часто используются методы седиментационного анализа, гель-хроматография и гель-электрофорез. Определение молекулярной массы белков методами седиментационного анализа проводят в ультрацентрифугах , в которых удается создать центробежные ускорения (g), превышающие в 200000 и более раз ускорение земного притяжения. Обычно вычисляют молекулярную массу по скорости седиментации молекул белка или седиментационному равновесию. По мере перемещения молекул от центра к периферии образуется резкая граница растворитель-белок (регистрируется автоматически). Оптические свойства растворителя и белка используются при определении скорости седиментации; последнюю выражают через константу седиментации s, которая зависит как от массы, так и от формы белковой частицы:
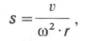
где v – скорость перемещения границы растворитель-белок, см/с; ω – угловая скорость ротора, рад/с; r – расстояние от центра ротора до середины ячейки с раствором белка, см. Константа седиментации имеет размерность времени (ее выражают в секундах). Величина константы седиментации, равная 1•10–13 с, условно принята за единицу и названа сведбергом (S). Значения констант седиментации большинства белков лежат в пределах 1–50 S, хотя в ряде случаев эти значения превышают 100 S.
Для вычисления молекулярной массы (М), помимо константы седиментации, необходимы дополнительные сведения о плотности растворителя и белка и другие согласно уравнению Сведберга:
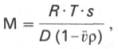
где R – газовая постоянная, эрг/(моль•град); Т – абсолютная температура (по шкале Кельвина); s – константа седиментации; ρ – плотность растворителя; v – парциальный удельный объем молекулы белка; D - коэффициент диффузии.
Определение молекулярной массы белков методом ультрацентрифугирования требует много времени и сложной и дорогостоящей аппаратуры. Поэтому в последние годы разработаны два более простых метода (гель-хроматография и электрофорез). При использовании гель-хроматографии в первую очередь требуется откалибровать колонку. Для этого через колонку с сефадексом пропускают несколько белков с известными молекулярными массами и строят график, откладывая значения логарифмов молекулярной массы против их элюционных объемов, которые находят, как показано на рис. 1.9.
Известно, что между логарифмом молекулярной массы белка, имеющего сферическую форму, и элюционным объемом существует прямая зависимость. Поэтому легко определить молекулярную массу исследуемого белка, зная его объем элюции. Второй разновидностью этого метода является тонкослойная гель-хроматография. Длина пробега белка (в миллиметрах) через тонкий слой сефадекса находится в логарифмической зависимости от молекулярной массы белка (рис. 1.10).
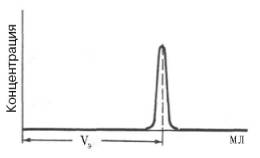
Рис. 1.9. Измерение объема элюции (VЭ).
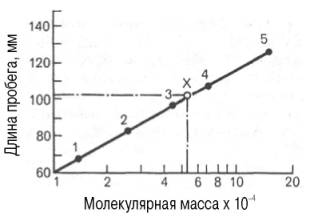
Рис. 1.10. Зависимость между длиной пробега белковых частиц при гель-хроматографии в тонком слое сефадекса Г-150 (сверхтонкого) и их молекулярными массами (в полулогарифмической системе координат).
1 - рибонуклеаза; 2 - химотрипсиноген; 3 -яичный альбумин; 4 - сывороточный альбумин; 5 - γ-глобулин; Х - белок с неизвестной молекулярной массой.
Гель-хроматография, кроме простоты и быстроты, имеет дополнительное преимущество: не требуется выделять белок в чистом виде, так как примеси других белков не мешают определению, поскольку каждый из них проходит через колонку со свойственной ему скоростью, определяемой молекулярной массой. Это обстоятельство широко используется в энзимологии, когда оказывается возможным определение молекулярной массы даже очень небольшого количества фермента в присутствии других белков, не обладающих аналогичной каталитической активностью.
При использовании диск-электрофореза в полиакриламидном геле для определения молекулярной массы белков также строят график зависимости между логарифмом молекулярной массы калибровочных белков и подвижностью белковых частиц в полиакриламидном геле, а затем, определив подвижность исследуемого белка, по графику находят его массу (рис. 1.11). Электрофорез проводят в присутствии детергента додецилсульфата натрия, так как только в этом случае наблюдается прямая пропорциональная зависимость между молекулярной массой и подвижностью белков. Белки с четвертичной структурой при этих условиях распадаются на субъединицы, поэтому метод находит широкое применение для определения молекулярной массы субъединиц белка.

Рис. 1.11. Зависимость между молекулярной массой и относительной подвижностью белка при диск-электрофорезе в полиакриламидном геле в присутствии додецилсульфата натрия (в полулогарифмической системе координат).
1 - сывороточный альбумин; 2 - яичный альбумин; 3 - пепсин; 4 - химотрипсиноген; 5 - мио-глобин; 6 - цитохром с; Х - белок с неизвестной молекулярной массой.
Недавно предложен новый масс-спектрометрический метод (так называемый лазерный десорбционно-ионизационный метод), позволяющий определять молекулярную массу небольших пептидов (вазопрессин, инсулин) и крупных биополимерных молекул и, кроме того, структуру биомолекул.