Липопротеины
ЛИПОПРОТЕИНЫ (липопротеиды), комплексы, состоящие из белков (аполипопротеинов; сокращенно - апо-липопротеины) и липидов, связь между к-рыми осуществляется посредством гидрофобных и электростатич. взаимодействий. Липопротеины подразделяют на свободные, или р-римые в воде (липопротеины плазмы крови, молока, желтка яиц и др.), и нерастворимые, т. наз. структурные (липопротеины мембран клетки, миелиновой оболочки нервных волокон, хлоропластов растений). Нековалентная связь в липопротеинах между белками и липидами имеет важное биол. значение. Она обусловливает возможность своб. обмена липидов и модуляцию св-в липопротеинов в организме. Среди своб. липопротеинов (они занимают ключевое положение в транспорте и метаболизме липидов) наиб. изучены липопротеины плазмы крови, к-рые классифицируют по их плотности. Чем выше содержание в них липидов, тем ниже плотность липопротеинов (табл. 1). Различают липопротеины очень низкой плотности (ЛОНП), низкой плотности (ЛНП), высокой плотности (ЛВП) и хиломикроны. Каждая группа липопротеинов очень неоднородна по размерам частиц (самые крупные - хиломикроны) и содержанию в ней апо-липопротеинов. Все группы липопротеинов плазмы содержат полярные и неполярные липиды в разных соотношениях.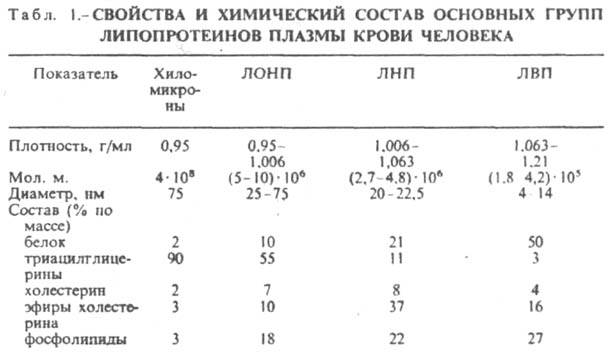
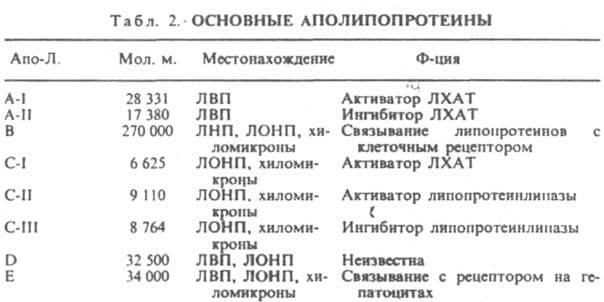
Выделяют липопротеины плазмы крови обычно методом флотации, используя водные р-ры бромидов Na или К разл. плотности. К структурным липопротеинам относятся комплексы липидзависимых ферментов (напр., аденилатциклаза), рецспторные комплексы (напр., рецептор ацетилхолина), к-рые входят в состав мембран биологических. Их строение и биол. св-ва очень разнообразны.
Литература
Климов А. Н., в сб.: Биохимия липидов и их роль в обмене веществ, М., 1981, с. 45-75; Structural and functrond aspects of lipoproteins in living systems, ed. by E. Tria, A. M. Scanu, L. N. Y., 1969. Н. В. Проказова.
Синонимы/примеры:*
Липопротеины высокой плотности; Липопротеины низкой плотности; Липопротеины очень низкой плотности; Хиломикроны
*подобраны ИИ, возможны неточности
Ещё по теме
Липопротеины — строение и функции
Липопротеины — строение и функции
Липопротеины плазмы крови — строение, классификация и роль в организме
Липопротеины — функции и значение в организме
Липиды — классификация и функции
Липиды — свойства и классификация
Структура и функции биологических мембран
Белки плазмы крови — функции и разделение на фракции
Нарушения липидного обмена и их последствия
Печень и липидный обмен — ключевые процессы и функции
Белки плазмы крови — функции и значение
Гликопротеины — структура, функции и биологическая роль
Типы белков и их функции