Регуляция энергетического обмена
Биохимический процесс усвоения пищи и образования АТФ должны постоянно приспосабливаться к изменению энергетических потребностей клеток. Необходимость согласования производства и потребления АТФ следует уже из того факта, что суммарное содержание коферментов в организме незначительно. Калорийность суточного рациона человека составляет примерно 12000 кДж. При к.п.д. 50% такая энергия достаточна для образования 120 молей АТФ, т. е. примерно 65 кг. Однако в организме человека содержится всего 3-4 г свободных адениновых нуклеотидов (АМФ, АДФ и АТФ). Следовательно, каждая молекула АДФ должна ежедневно тысячекратно фосфорилироваться в АТФ и вновь дефосфорилироваться.
А. Дыхательный контроль
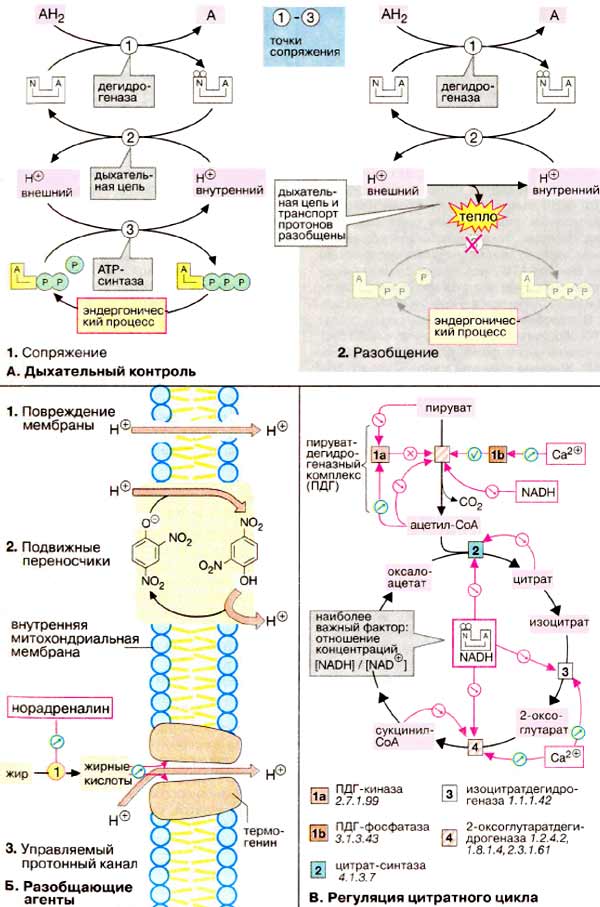
Простой механизм регуляции образования и потребления АТФ (АТР) называется дыхательным контролем. Он основан на сопряжении упомянутых процессов с общими коферментами и другими факторами (на схеме слева). Если клетка не расходует АТФ, едва ли в митохондриях имеется АДФ. В отсутствие АДФ АТФ-синтаза (3) не в состоянии использовать протонный градиент на внутренней митохондриальной мембране. Это в свою очередь тормозит электронный перенос вдыхательной цепи (2), вследствие чего НАДН не может быть вновь окислен в НАД+. Возникающее в результате высокое соотношение НАДН/НАД+ тормозит цитратный цикл (схема В) и замедляет тем самым потребление субстрата АН2 (1). И наоборот, высокие скорости потребления АТФ стимулируют усвоение пищи и дыхательную цепь по тому же механизму.
Если создание протонного градиента (на схеме справа) подавлено, процессы окисления субстрата (1) и переноса электронов (2) протекают значительно быстрее, чем обычно. При этом вместо синтеза АТФ выделяется тепло.
Б. Разобщающие агенты
Вещества, которые функционально разделяют между собой окисление и фосфорилирование, называются разобщающими агентами. Они содействуют переносу протонов из межмембранного пространства в матрикс без участия АТФ-синтазы. Разобщение может возникать, например, в результате механического повреждения внутренней мембраны (1) или действия таких веществ, как 2,4-динитрофенол (2), являющихся переносчиками протонов через мембрану. Природным разобщающим агентом является термогенин (3), протонный канал (см. с. 216) в митохондриях бурых жировых клеток. Бурый жир обнаружен у новорожденных и животных, впадающих в зимнюю спячку и служит для теплообразования. При охлаждении организма норадреналин активирует гормонзависимую липазу. Благодаря интенсивному липолизу в организме образуется большое количество свободных жирных кислот, которые распадаются в результате β-окисления и в дыхательной цепи. Так как жирные кислоты одновременно открывают протонный канал термогенина, их распад не зависит от наличия АДФ, т. е. протекает с максимальной скоростью и генерирует энергию в форме тепла.
В. Регуляция цитратного цикла
Самым важным фактором регуляции цикла является отношение НАДН/НАД+ (NADH/NAD+). НАДН наряду с пируватдегидрогеназой (ПДГ) и оксоглутаратдегидрогеназой (ОГД, см. с. 136) ингибирует также цитрат-синтазу и изоцитрат-дегидрогеназу. За исключением изоцитратдегидрогеназы упомянутый ферменты также ингибируются конечным продуктом реакции ацетил- и соответственно сукцинил-КоА или цитратом. Активность ферментов регулируется также процессом взаимопревращения. На схеме эти процессы представлены на примере пируватдегидрогеназного комплекса. Инактивированная протеинкиназа [1а] ингибируется субстратом (пируватом) и активируется продуктом реакции ацетил-КоА. Соответствующая протеинфосфатаза [1б] активируется ионами Ca2+, так же как изоцитратдегидрогеназа [3] и оксоглутаратдегидрогеназный комплекс [4], что особенно важно для процесса мышечного сокращения.