Дыхание
ДЫХАНИЕ, совокупность процессов, обеспечивающих поступление в организм атмосферного или растворенного в воде О2, использование его в окислит.-восстановит. р-циях, а также удаление из организма СО2 и нек-рых др. соед. - конечных продуктов обмена в-в. Играет фундам. роль в энергообеспечении и метаболизме у большинства организмов. При дыхании кислород участвует гл. обр. в окислении орг. соед. с образованием Н2О или Н2О2 (в нек-рых случаях - О2-) или включается в молекулу окисляемого в-ва. Нек-рые организмы (гл. обр. мн. бактерии) могут использовать в качестве акцептора электронов не только О2, но и др. соед. с высоким сродством к электрону, напр., нитраты и сульфаты. В этих случаях иногда говорят о "нитратном" и "сульфатном" дыхании в отличие от аэробного (кислородного) дыхания. У высших организмов дыхание - сложный комплекс физиол. и биохим. процессов, в к-ром можно выделить ряд осн. стадий: 1) внеш. дыхание поступление О2 из среды в организм, осуществляемое с помощью спец. органов дыхания (легких, жабр, трахей и т.д.) или через пов-сть тела (напр., у кишечно-полостных); 2) транспорт О2 от органов дыхания ко всем др. органам, тканям и клеткам у большинства животных эта ф-ция обеспечивается кровеносной системой при участии спец. белков - переносчиков кислорода (гемоглобин, миоглобин, гемоцианин и др.); 3) тканевое, или клеточное, дыхание - собственно биохим. процесс восстановления О2 в клетках при участии большого числа разных ферментов. Дыхание многих, в первую очередь одноклеточных, организмов сводится к клеточному дыханию, а стадии 1 и 2 обеспечиваются диффузией О2. В клеточном дыхании осн. часть потребления О2 аэробными организмами (их на Земле абс. большинство) связана с обеспечением клетки энергией в процессе окислительного фосфорилирования, к-рый у животных и растений осуществляется в спец. субклеточных структурах - митохондриях. В окружающей митохондрию бислойной фосфолипидной мембране находится система окислит.-восстановит. ферментов, наз. дыхательной или электронотранспортной цепью (см. рис.). Эта цепь катализирует перенос электронов (протонов) от ряда продуктов обмена в-в (т. наз. субстраты окисления) к О2. Окислит.-восстановит. потенциал субстратов окисления колеблется, как правило, от - 0,4 до 0 В. Наиб. важные субстраты окисления - в-ва, образующиеся при функционировании цикла трикарбоновых к-т (напр., янтарная к-та, восстановленный кофермент никотинамидадениндинуклеотидфосфат, жирные к-ты, нек-рые аминокислоты и продукты метаболизма углеводов). Б. ч. своб. энергии переноса электронов в дыхат. цепи трансформируется первоначально в энергию разности электрохим. потенциалов ионов Н+ (D m Н) на мембране митохондрий, к-рая далее используется для термодинамически невыгодного синтеза АТФ из аденозиндифосфата и неорг. фосфата при окислит. фосфорилировании.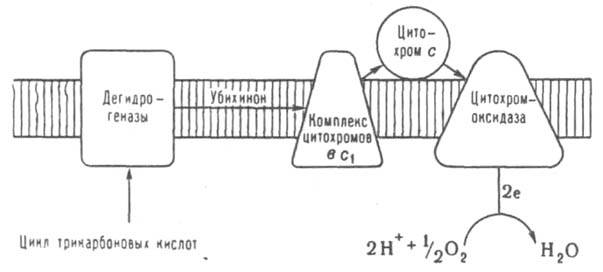
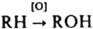 , напр., гидроксилирование стероидов; второй атом О в молекуле О2 восстанавливается при этом до Н2О). Эта электронотранспортная цепь особенно активна в печени животных. В биохимии клеточного дыхания различают неск. осн. р-ций с участием О2: 1) катализируемое оксидазами ("аэробными гидрогеназами") четырехэлектронное восстановление О2 до Н2О или двухэлектронное до Н2О2:
, напр., гидроксилирование стероидов; второй атом О в молекуле О2 восстанавливается при этом до Н2О). Эта электронотранспортная цепь особенно активна в печени животных. В биохимии клеточного дыхания различают неск. осн. р-ций с участием О2: 1) катализируемое оксидазами ("аэробными гидрогеназами") четырехэлектронное восстановление О2 до Н2О или двухэлектронное до Н2О2: 2RH2 + O2 : 2R + 2Н2O; RH2 + O2 : R + H2O2
2) Включение обоих атомов О2 в молекулу окисляемого в-ва, катализируемое диоксигеназами (оксигеназами):
RH2 + O2 : R(OH)2
3) Включение одного из атомов О2 в молекулу окисляемого в-ва, др. атом О восстанавливается с образованием Н2О в результате окисления второго субстрата:
RH + R' Н2 + О2 : ROH + R' + Н2О
Ферменты, катализирующие эту р-цию, - монооксигеназы. В состав активных центров ферментов, взаимодействующих с О2, обычно входят ионы переходных металлов (медь, гемовое или негемовое железо) или флавины (коферментные формы витамина рибофлавина). Интенсивность дыхания организмов, тканей, клеток принято выражать в кол-ве О2, потребляемого за единицу времени на единицу массы (напр., в мг О2.мин-1 г-1). Важный показатель интенсивности дыхания высших позвоночных - кол-во воздуха, вентилируемого легкими в 1 мин (наз. минутным объемом дыхания, или МОД). Эти величины служат важнейшим показателем уровня энергетич. обмена организма. У человека МОД в состоянии покоя составляет 5-8 л/мин, во время физич. работы - до 100 и более л/мин. Соед., подавляющие дыхание (дыхат. яды), выключают энергообеспечение организма и потому являются быстродействующими ядами. Классич. дыхат. яды (цианиды, изоцианиды, сульфиды, азиды, СО и NO) угнетают концевой фермент дыхат. цепи митохондрий (цитохром-с-оксидазу). Эти же соед. угнетают транспорт О2 по организму, связываясь с гемоглобином. Др. важный класс дыхат. ядов - гидрофобные орг. в-ва, часто хиноидной природы, выступающие как антагонисты убихинона (замещенного 1,4-бензохинона), играющего ключевую роль во мн. стадиях переноса электронов по дыхат. цепи. Сильнейшие яды этого класса - токсич. антибиотики (ротенон, пирицидин, антимицин, миксотиазол), 2-гептил-4-гидроксихинолин-N-оксид; их используют в исследованиях тканевого дыхания. Способность к умеренному подавлению убихинон-зависимых р-ций в дыхат. цепи свойственна мн. лек. ср-вам (напр., барбитуратам), фунгицидам и пестицидам. Лит. Рэкер Э., Биоэнергетические механизмы: новые взгляды, пер. с англ., М.. 1979; Мецлер Д., Биохимия, пер. с англ.. т 2. М.. 1980. с. 361 445: Константинов А. С., Общая гидробиология, 4 изд., М., 1986. гл. 6; Скулачев В. П., Энергетика биологических мембран. М.. 1988; Molecular mechanisms of oxygen activation, N.Y., 1974; Wikstrdm M., Saraste M., The mitochondrial respiratory chain, в сб. Bioenergetics. Amst, 1984. p. 49-94. А. А. Константинов.