Дыхание и брожение
А. Аэробное и анаэробное окисление глюкозы
В присутствии кислорода (в аэробных условиях) большинство клеток животных получают энергию за счет полного разрушения питательных веществ (липидов, аминокислот и углеводов), т. е. за счет окислительных процессов. В отсутствие кислорода (анаэробные условия) клетка может синтезировать АТФ (АТР) только за счет гликолитического разрушения глюкозы. Хотя такое разрушение глюкозы, заканчивающееся образованием лактата, дает незначительную энергию для синтеза АТФ, этот процесс имеет решающее значение для существования клеток при недостатке или в отсутствие кислорода.
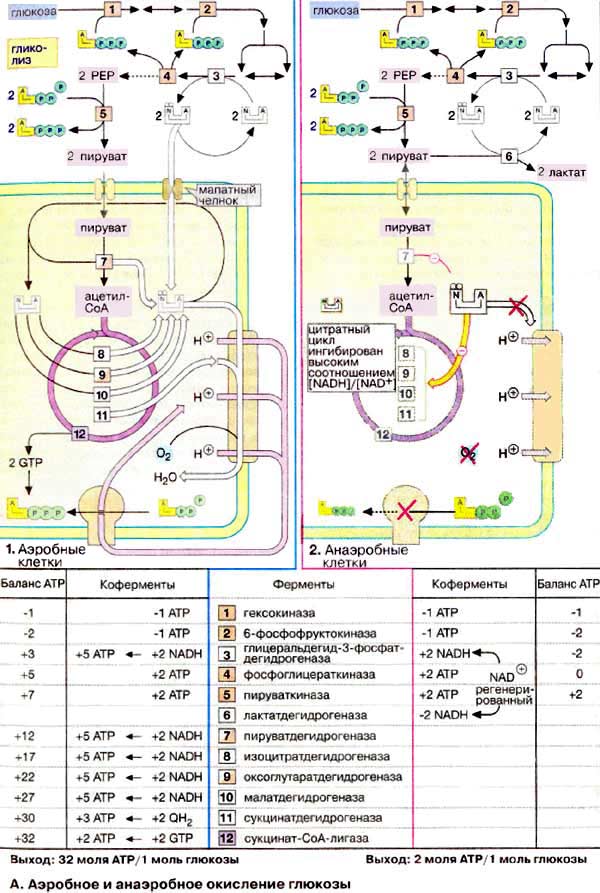
В аэробных условиях (на схеме слева) АТФ образуется почти исключительно за счет окислительного фосфорилирования. Жирные кислоты в виде ацилкарнитина попадают в матрикс митохондрий, где подвергаются β-окислению с образованием ацил-КоА. Глюкоза в цитоплазме превращается в пируват путем гликолиза. Пируват транспортируется в митохондриальный матрикс, где декарбоксилируется пируватдегидрогеназным комплексом с образованием ацетил-КоА. Восстановительные эквиваленты [2 НАДН + Н+ (NADH + Н+) на молекулу глюкозы], высвобождающиеся при гликолизе, переносятся в матрикс митохондрий малатным челноком. Образующиеся из жирных кислот ацетильные остатки окисляются до CO2 в цитратном цикле. Деградация аминокислот также приводит к ацетильным остаткам или продуктам, которые непосредственно включаются в цитратный цикл. В соответствии с энергетическими потребностями клетки восстановительные эквиваленты переносятся дыхательной цепью на кислород. При этом высвобождается химическая энергия, которая путем создания протонного градиента используется для синтеза АТФ.
В отсутствие кислорода, т. е. в анаэробных условиях (на схеме справа), картина полностью меняется. Так как электронных акцепторов для дыхательной цепи не хватает, НАДН + Н+ и QH2 не могут окисляться повторно. Вследствие этого останавливается не только митохондриальный синтез АТФ, но почти весь обмен веществ в митохондриальном матриксе. Главной причиной такой остановки является высокая концентрация НАДН (NADH), ингибирующая цитратный цикл и пируватдегидрогеназу. Останавливаются также процесс β-окисления и функционирование малатного челнока, зависящие от наличия свободного НАД+. Поскольку энергия уже не может быть получена за счет деградации аминокислот, клетка становится полностью зависимой в энергетическом отношении от потребления глюкозы при гликолизе. При этом обязательным условием является постоянное окисление образующегося НАДН + Н+. Так как этот процесс уже не может идти в митохондриях, в клетках животных, функционирующих в анаэробных условиях, пируват восстанавливается до лактата, который поступает в кровь. Процессы этого типа называют брожением. Продукция АТФ при этих процессах незначительна: при образовании лактата возникают только 2 молекулы АТФ на молекулу глюкозы.
Для того чтобы оценить число образованных в аэробном состоянии молекул АТФ, необходимо знать так называемое Р/О-соотношение, т. е. молярное соотношение синтезированных АТФ (Р) и воды (О). Во время переноса двух электронов от НАДН на О2 в межмембранное пространство транспортируются около 10 протонов и только 6 молекул убихинола (QH2). Для синтеза АТФ АТФ-синтаза нуждается в трех ионах Н+, так что максимальное возможное P/О-соотношение составляет примерно 3 или, соответственно, 2 (для убихинола). Нужно, однако, учитывать, что при переходе метаболитов в матрикс и обмене митохондриального АТФ4- на цитоплазматический АДФ3- в межмембранном пространстве также расходуются протоны. Поэтому при окислении НАДН P/О-соотношение скорее всего составляет 2,5, а при окислении QH2 — 1,5. Если на основе этих величин рассчитать энергобаланс аэробного гликолиза, получается, что окисление одной молекулы глюкозы сопровождается синтезом 32 молекул АТФ.