Деградация аминокислот
В обзоре представлены многочисленные пути метаболизма аминокислот. Дополнительные подробности приведены на сс. 402 и 403.
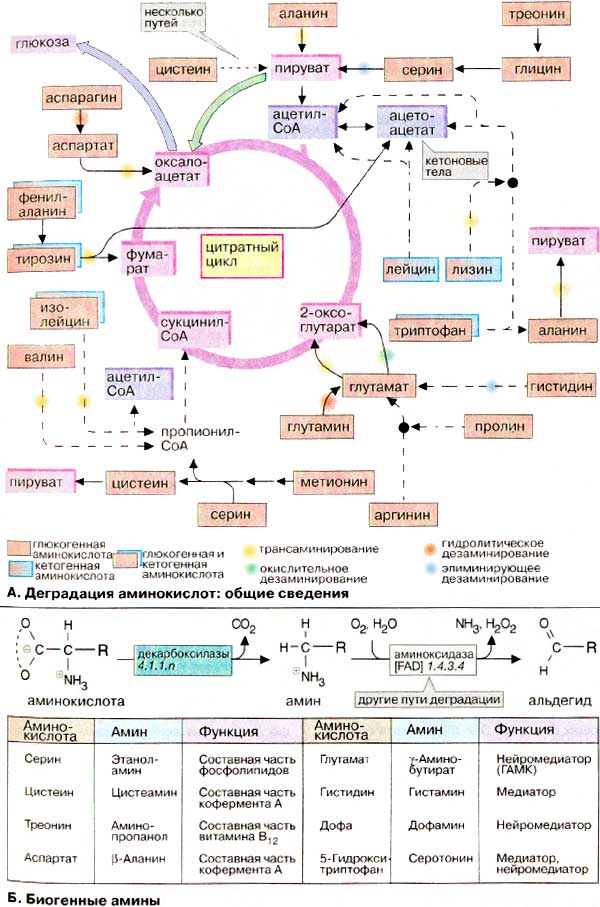
А. Деградация аминокислот: общие сведения
Углеродные скелеты 20 белковых аминокислот превращаются в итоге в семь различных продуктов деградации (на схеме окрашены в розовый и светло-голубой цвета). Пять метаболитов (2-оксоглутарат, сукцинил-КоА, фумарат, оксалоацетат и пируват) служат предшественниками в процессе глюконеогенеза. Первые четыре являются еще и промежуточными продуктами цитратного цикла, в то время как пируват может быть переведен пируватдекарбоксилазой в оксалоацетат и тем самым стать участником глюконеогенеза (зеленая стрелка). Аминокислоты, деградация которых поставляет один из пяти упомянутых метаболитов, называются глюкогенными аминокислотами. За двумя исключениями (лизин и лейцин, см. ниже) глюкогенными являются все белковые аминокислоты.
Два других продукта распада (ацетоацетат и ацетил-КоА) не могут включаться в глюконеогенез в организме животных. Они используются для синтеза кетоновых тел, жирных кислот и изопреноидов (см. сс. 174, 304). Поэтому аминокислоты, которые разрушаются с образованием ацетил-КоА или ацетоацетата, называются кетогенными аминокислотами. Фактически кетогенными являются только лейцин и лизин. Некоторые аминокислоты поставляют продукты деградации, являющиеся глюкогенами и кетогенами. К этой группе принадлежат фенилаланин, тирозин, триптофан и изолейцин.
Существует несколько путей удаления аминогруппы во время распада аминокислоты (дезаминирования). Обычно NH2-группа переносится путем трансаминирования на 2-оксоглутарат. Образующийся глутамат в дальнейшем вновь превращается в 2-оксоглутарат с помощью глутаматдегидрогеназы (окислительное дезаминирование, зеленая метка). В этой реакции образуется свободный аммиак (NH3), который у высших животных превращается в мочевину и выводится из организма. Аммиак освобождается также при гидролизе амидных групп аспарагина и глутамина (гидролитическое дезаминирование, оранжевая метка). Другим превращением, при котором образуется NH3, является элиминирующее дезаминирование серина в пируват (голубая метка, см. с. 402).
Б. Биогенные амины
Моноамины, так называемые биогенные амины, образуются при декарбоксилирование аминокислот. Некоторые из этих соединений являются составными частями других биомолекул. Так. в состав фосфолипидов кроме аминокислоты серина может входить соответствующий биогенный амин этаноламин. Цистеамин и β-аланин являются структурными элементами кофермента А и пантетеина. Образованный из треонина аминопропанол является структурным элементом витамина B12.
Некоторые биогенные амины действуют как сигнальные вещества. Важным нейромедиатором является образующаяся из глутамата γ-аминомасляная кислота [ГАМК (GABA), см. с. 338]. Другие нейромедиаторы образуются путем декарбоксилирования небелковых аминокислот. Так, из 3,4-дигидроксифенилаланина (дофа) образуется медиатор дофамин. Дофамин является одновременно предшественником катехоламинов адреналина и норадреналина. Нарушения метаболизма дофамина служат причиной болезни Паркинсона. Из триптофана через промежуточный 5-гидрокситриптофан образуется серотонин, соединение с широким спектром действием.
Многие моноамины и катехоламины инактивируются аминоксидазой (моноаминоксидазой, "МАО") путем дезаминирования с одновременным окислением в альдегиды. Следовательно, ингибиторы МАО играют важную роль при фармакологическом воздействии на метаболизм нейромедиаторов.