Мутагeны
МУТАГEНЫ (от мутации и греч. -genes-рождающий, рожденный), хим. и физ. факторы, вызывающие наследств. изменения - мутации. Впервые искусств. мутации получены в 1925 Г. А. Надсе-ном и Г. С. Филипповым у дрожжей действием радиоактивного излучения радия; в 1927 Г. Мёллер получил мутации у дрозофилы действием рентгеновских лучей. Способность хим. в-в вызывать мутации (действием иода на дрозофилы) открыта в 1932 В. В. Сахаровым.
Мутагены химические. Различают мутагены прямого действия -соед., реакц. способность к-рых достаточна для хим. модификации ДНК, РНК и нек-рых белков, и промутагены - в-ва, к-рые сами по себе инертны, но превращ. в организме в мутагены (в осн. в результате ферментативного окисления системой микро-сомных многоцелевых оксидаз). Последние часто называют "конечными" мутагенами. Так, немутагeнный 1,2-бензопкрен (бен-зо[а]пирен) в организме окисляется до 7,8-дигидрокси-9,10-эпокси-7,8,9,10-тетрагидро-1,2-бензопирена, один из стерео-изомеров к-рого обладает мутагенной, а также канцерогенной активностью:
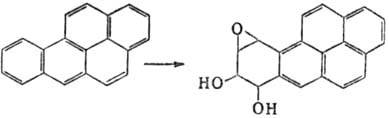
Др. пример - немутагенный N-нитрозодиметиламин, к-рый в печени подвергается ферментативному окислит. деметили-рованию и превращ. в высокомутагенный и канцерогенный метилдиазогидроксид (см. Канцерогенные вещества).
Мишенью действия мутагенов в клетке являются гл. обр. ДНК и, возможно, нек-рые белки. К последним относят в осн. белки, играющие структурную роль в организации генома или принимающие участие в репликации (самовоспроизведении молекулы нуклеиновых к-т), рекомбинации (перераспределении генетич. материала родителей в потомстве) или репарации (восстановлении поврежденной структуры ДНК).
Среди мутагенов наиб. обширен класс электроф. алкилирующих мутагенов, к к-рым относят не только типичные алкилирующие агенты (диазоалканы, эфиры серной к-ты и алкансульфо-кислот), но и эфиры фосфорной и азотной к-т, аминоэтили-рующие реагенты (2-хлорэтиламин, этиленимин и их производные), оксиэтилирующие агенты (этиленоксид и его производные) и альдегиды. К этому же классу мутагенов относят N-нитрозо-N-алкиламиды карбоновых к-т, N-нитрозо-N-алкилуретаны, N-нитрозо-N-алкилмочевины, N-алкил-N-нитрозо-N'-нитрогуанидины, являющиеся, по-видимому, наиб. активными из известных мутагенов. Эти соед. сами по себе лишены алкилирующих св-в, но при их гидролитич. распаде образуются активные алкилдиазогидрохсиды (иногда считают, что образуются своб. алкилкарбкатионы, что менее вероятно).
Электроф. реагентами являются также подавляющее большинство конечных мутагенов, образующихся из синтетич. и прир. в-в. Пример последних - глюкозид циказин (b-D-глю-козид метилазоксиметанола), к-рый под действием ферментов трансформируется в метилдиазогидроксид.
Механизм мутагенного действия простейших алкилирующих агентов довольно хорошо изучен. Характер повреждений ДНК при воздействии этих агентов м.б. предсказан с помощью ф-лы Свена-Скотта:
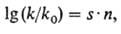
где k- константа скорости бимолекулярной р-ции алкили-рующего агента с нуклеофилом; k0 - константа скорости бимолекулярной р-ции алкилирующего агента с водой, выбранной в качестве стандарта; n-константа, характеризующая нуклеофильность субстрата; .s-мера чувствительности скорости р-ции к изменению п.
Из ф-лы следует, что при алкилировании биополимеров, имеющих неск. нуклеоф. центров, доля продуктов алкили-рования центра с низкой нуклеофильностью должна быть выше при действии агента с низкой константой s.
Наибю нуклеоф. центр в молекуле ДНК-положение 7 в гуанине. Поэтому диметилсульфат и метилметансульфоиат (s соотв. 0,9 и 0,86) алкилируют ДНК в осн. по этому положению (см. ф-лу I; везде R-цепь ДНК), а выход продуктов алкилирования центров с меньшей нуклеофильностыо мал. При действии на ДНК этилметансульфоната и N-нитрозо-N-метилмочевины (s соотв. 0,67 и 0,42) доля 7-алкилгуанина в общем выходе продуктов алкилирования снижается и возрастает вклад продуктов алкилирования центров с низкой нуклеофильностью - межнуклеотидных фосфатных групп и атомов кислорода в основаниях. При алкилировании ДНК N-нитрозо-N-этилмочевиной (s ок. 0,26) осн. продукт р-ции -алкилир. остатки фосфорной к-ты и О-алкилир. основания: О6-этилгуанин (II), О2-этилцитозин (III), О2- и О4-этилтимины (соотв. IV и V).
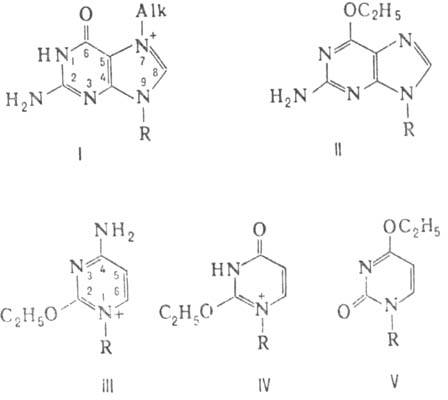
Синтез новой нити ДНК на ДНК-матрице, содержащей 7-алкилгуаниновые звенья, проходит без ошибок и мутаций не возникает. Напротив, полинуклеотид, синтезированный на матрице, содержащей О-алкилир. звенья, содержит ошибочно включенные пуриновые и пиримидиновые основания. С возрастанием способности к О-алкилированию у простейших алкилирующих агентов возрастает канцерогенная активность.
Мутации, возникающие при действии метилметансульфо-ната, не алкилирующего основания ДНК по атому О, являются следствием ошибок работы ферментов клетки, восстанавливающих исходное (неизмененное) состояние ДНК.
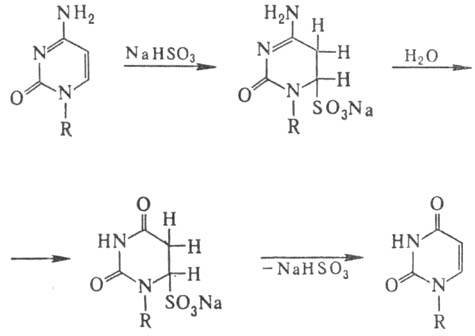
Спектр повреждений ДНК при действии нуклеоф. мутаге-нов (гидроксиламин, О-метилгидроксиламин, гидразины, бисульфит Na) значительно уже. В осн. это модификация цитозина, направление и механизм к-рой иллюстрирует след. схема:
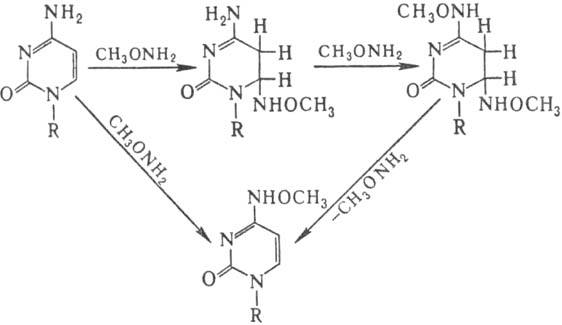
Др. нуклеоф. агенты реагируют аналогично, но константы скорости отдельных стадий могут существенно меняться. Особенность действия бисульфита на ДНК-замена цитози-новых звеньев на урацилъные в результате дезаиминирования цитозина по схеме:
Ряд мутагенов вызывают мутации, не связываясь ковалентно с ДНК. Так, нек-рые гетероциклич. соед. (напр., акридин и его производные), обладающие жесткой плоской структурой молекулы, встраиваются (интеркалируют) между смежными, расположенными стопкой, парами оснований двойной спирали ДНК. В этом случае матричный синтез на ДНК протекает с ошибками. В синтезируемой нити ДНК оказывается на один нуклеотид больше или меньше обычного и возникают мутации.
Особый класс мутагенов составляют соед., представляющие со-бой аналоги оснований ДНК-5-галогенурацилы, 2-амино-и 6-метиламинопурины и др. Галогенурацилы включаются в ДНК при матричном синтезе вместо тимина, 2-амино-пурин-вместо аденина. Вследствие различий в положении кетоенольного равновесия у тимина и галогенурацилов (при включении последних в ДНК) увеличивается частота ошибочных спариваний оснований и возникают ошибки при репликации.
Существуют также мутагены, ингибирующие синтез предшественников ДНК. Мишень таких мутагенов-ферменты, синтезирующие компоненты ДНК. Считается, что в результате подавления синтеза предшественников происходит замедление или даже остановка синтеза ДНК. В этих условиях повышается вероятность того, что решшкац. система клетки может пропустить отсутствующий нуклеотид, либо включить вместо отсутствующего нуклеотида другой, ошибочный; следствие обоих событий-мутация.
Мутагенные и канцерогенные св-ва хим. в-в тесно связаны между собой. Поэтому выявление возможных мутагенов в окружающей среде, испытание на мутагенность продуктов пром. синтеза (красители, лек. ср-ва, пестициды и др.)-важная задача совр. генетики. Разработаны тест-системы для экспресс-обнаружения мутагенов. Наиб. часто используют тест Б. Эймса и его модификации. Для их осуществления используют специально полученные штаммы бактерий Salmonella typhimurium, к-рые не способны синтезировать гистидин из-за генетич. нарушений. Этот штамм поэтому не может расти в среде, в к-рой отсутствует эта аминокислота. Др. его особенность-способность в результате обратной мутации приобретать исходную способность синтезировать гистидин из обычных предшественников (NH3 и др.). Частота обратных мутаций заметно увеличивается под действием мутагенов и может служить критерием их активности. В тестах используют лишенную гистидина питат. среду, в к-рую добавляют экстракт из печени крыс, содержащий ферменты эндоплаз-матич. ретикулума, способные превращать в-во в мутаген-ную (канцерогенную) форму. В случаях, когда в-во обладает мутагенной активностью, наблюдается активный рост колоний бактерий. Время тестирования-ок. 24 ч (на тестирование с использованием эксперим. животных затрачивается 2-3 года).
Мутагены физические. Мутации при действии физических мутагенов возникают так же, как и при действии мутагенов химических. Вначале возникает первичное повреждение ДНК. Если оно не будет полностью исправлено в результате репарации, то при послед. репликативном синтезе ДНК будут возникать мутации. Специфика мутагенеза (процесса возникновения мутаций) при действии физ. факторов связана с характером первичных повреждений генома, вызываемых ими.
Подробно изучены повреждения ДНК, возникающие в результате действия электромагн. излучения разной длины волны. Электромагн. излучение с длиной волны больше 300 нм не поглощается ДНК, однако в нек-рых случаях может оказывать мутагенное действие, механизм к-рого заключается в поглощении кванта света молекулой сенсибилизатора и передачей энергии возбуждения на ДНК. При действии света с длиной волны 200-300 нм (lмакс поглощения ДНК 260 нм) происходит поглощение квантов света хромофорными группами ДНК (пуриновые и пиримидиновые основания) и переход последних в возбужденное состояние. В обоих случаях б.ч. поглощенной энергии рассеивается и основания ДНК возвращаются в исходное, невозбужденное, состояние, но часть возбужденных оснований подвергается фотохим. трансформации.
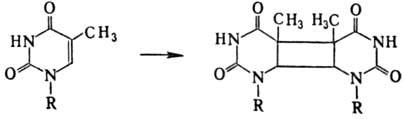
В наиб. степени это относится к тимину, остальные основания более устойчивы. Энергия излучения, поглощенная тимином, локализуется в осн. на двойной связи цикла; следствием этого является образование в составе ДНК димеров:
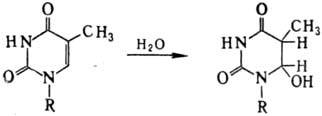
Возбужденная двойная связь способна также присоединять нуклеофилы с образованием, напр., нестойкого гидрата тимина:
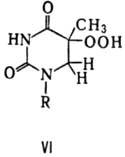
При облучении ДНК дальним УФ светом (185 нм) возможно и фотоокисление тимина с образованием его гидроперок-сида (ф-ла VI).
Предполагают, что осн. предмутац. повреждение ДНК, возникающее при действии УФ света, связано с образованием димеров тимина.
Электромагн. излучения еще более высокой энергии (рентгеновское и у-излучение) способны ионизовать в-во. Ионизация происходит случайным образом, поэтому молекулы, являющиеся наиб. распространенными в объекте, больше других под вергаются ионизации. При облучении живой материи, на 70-90% состоящей из воды, б. ч. энергии будет поглощена молекулами воды и поэтому мутагенный эффект при действии этих агентов возникает гл. обр. вследствие модификации ДНК продуктами радиолиза воды. Наиб. вклад в развитие радиац. поражения ДНК вносит радикал ОН.. При взаимод. с ДНК 80% всех радикалов ОН. атакуют основания ДНК, остальные-дезоксирибозную часть молекулы. Возникающие первичные продукты затем вступают в разнообразные вторичные р-ции как с теми же продуктами радиолиза воды, так и с кислородом, белками, низкомол. компонентами клетки, а также подвергаются диспропорционированию, изомеризации, гидролизу. Возникает широкий спектр разнообразных изменении первичной и вторичной структуры ДНК: измененные основания, апури-новые и апиримидиновые сайты (участки с удаленными основаниями), разрывы связей в дезоксирибозе, одно- и двунитевые разрывы цепей ДНК. Точная роль каждого из возникающих повреждений структуры ДНК в формировании мутагенного эффекта все еще остается невыясненной. Предполагают, что ключевую роль в этом процессе играют продукты радиолиза тимина.
В тимине первичной атаке радикалом ОН. подвергается двойная связь цикла с образованием двух радикалов с радикальными центрами в положениях 5 и 6, с преобладанием первого. В присут. кислорода из радикала образуется гидро-пероксид, к-рый уже при комнатной т-ре разлагается с раскрытием цикла:
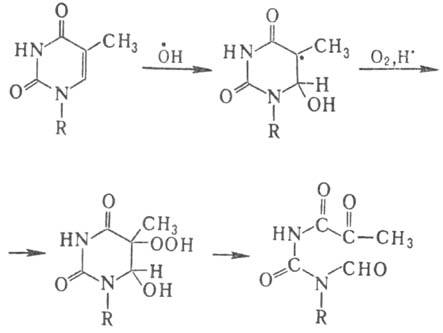
Радиационно-хим. изменения цитозина также протекают через стадию образования аналогичного, но еще более нестабильного гидропероксида. В случае цитозина и аденина возможно также дезаминирование оснований. Пуриновые основания (аденин, гуанин) реагируют с радикалом ОН. с меньшей скоростью. Идентифицированы, напр., продукт гидроксилирования аденина (8-гидроксиаденин), а также продукты раскрытия имидазольного кольца этих оснований.
В дезоксирибозном остатке молекулы ДНК радикал ОН. способен атаковать и отрывать атом Н из любого положения цикла. Конечный результат радиационно-хим. модификации этой части молекулы ДНК - образование малонового диальдегида, сопровождающееся разрывом N-гликозидной связи с отщеплением неизмененного основания, и, в итоге, однонитевых разрывов ДНК. Затем уже возникают и двунитевые разрывы ДНК в результате статистич. накопления однонитевых разрывов в противоположных участках двух цепей ДНК.
Известно прямое действие радиации на молекулы-мишени, в результате к-рого макромолекулы превращ. в своб. радикалы по схеме:
Судьба образовавшихся радикалов R., вероятно, схожа с судьбой радикалов, возникающих при непрямом действии радиации.
Механизм действия др. видов ионизирующего излучения (a-частицы, протоны, нейтроны, электроны, ускоренные ядра более тяжелых элементов) близок к механизму действия ионизирующего электромагн. излучения. Отличия обусловлены гл. обр. разницей в массе, заряде, энергии и глубине проникновения излучения в объект, способом ионизации макромолекул и др. Имеются сведения, что воздействие нек-рых др. физ. факторов, напр. звуковых колебаний, вибрации, могут также привести к мутации.
Мутагены применяют для получения штаммов микроорганизмов, продуцирующих в больших кол-вах антибиотики, аминокислоты, витамины и др. в-ва, чем прир. штаммы (такие штаммы используют в пром-сти), а также для выведения новых сортов растений.
Исп. литература для статьи «МУТАГEНЫ»: Первичные радиобиологические процессы, под ред. Н. В. Тимофеева-Ресовского, 2 изд., М., 1973; Рябченко Н.И., Радиация и ДНК, М., 1979; КогглД., Биологические эффекты радиации, пер. с англ., М., 1986; Рубин А.Б., ФрайкинГ.Я., "Успехи совр. биологии", 1987, т. 103, в. 3, с. 323-39; Chemical mutagens. Principles and methods for their detection, ed. by A. Hollander, v. 1-10, N.Y., 1971-86. A.M. Серебряный.