Рибонуклеиновые кислоты
Рибонуклеиновые кислоты [PHK(RNA)] представляют собой полимеры из нуклеозидфосфатных звеньев, соединенных фосфодиэфирной связью. В качестве азотистых оснований в РНК присутствуют урацил, цитозин, аденин и тимин. В РНК можно также встретить множество необычных и модифицированных азотистых оснований.
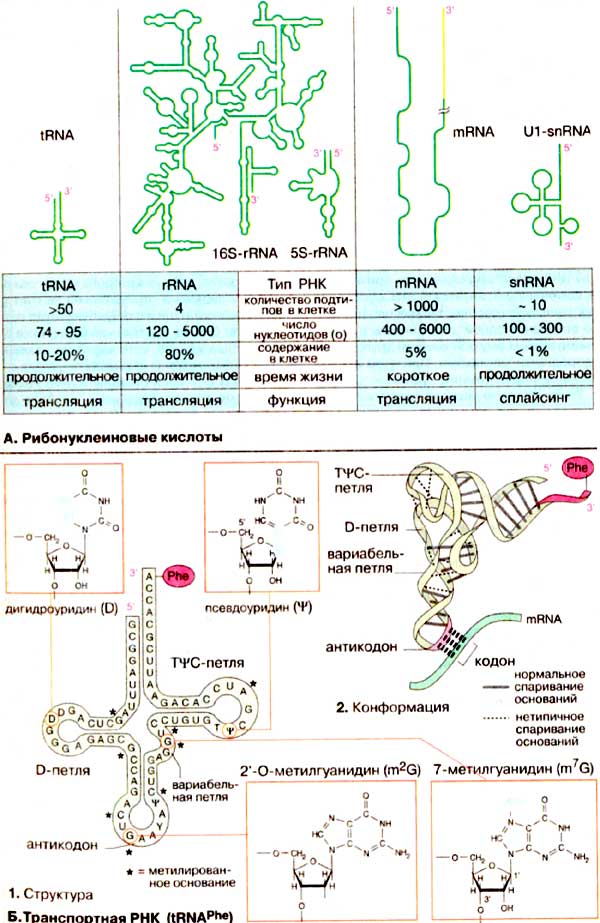
А. Рибонуклеиновые кислоты
РНК принимают участие во всех стадиях процесса генной экспрессии и биосинтеза белка. Свойства наиболее важных видов РНК приведены в таблице. Кроме того, здесь схематически показаны вторичные структуры молекул РНК.
В отличие от ДНК, РНК не образуют двойных спиралей, но содержат короткие участки со спаренными основаниями. Это приводит к образованию субструктур, которые при двумерном изображении напоминают «шпильки» и петли, образующие фигуру типа «кленового листа». В таких структурах двухцепочечные участки соединены петлями. Множество фрагментов, в которых чередуются структуры типа шпилька—петля, содержится в высокомолекулярных РНК, таких, например, как рибосомная 16S-рРНК (16S-rRNA) (в центре). Кроме того, эти фрагменты образуют трехмерные структуры; следовательно, РНК подобно белкам имеют четвертичную структуру. До настоящего времени установлена четвертичная структура небольших PHK, прежде всего тРНК (tRNA). Из иллюстраций, приведенных на схеме Б и на с. 93 очевидно, что трехмерная укладка структуры типа «кленовый лист» окончательно не установлена.
PHK клетки существенно различаются по размерам, строению и продолжительности существования. Преобладающую часть представляют рибосомные РНК [рРНК (rRNA)], которые в различных формах составляют структурный и функциональные части рибосом. Рибосомные РНК синтезируются в ядре в процессе транскрипции на ДНК, там же подвергаются процессингу и ассоциируют с рибосомными белками, образуя рибосому (см. сс. 210, 240). Приведенная на схеме А бактериальная 16S-рРНК, включающая 1542 нуклеотида, является компонентом малой рибосомной субчастицы, в то время как небольшая 5S-рРНК (из 120 нуклеотидов) входит в состав большой субчастицы.
Матричная РНК [мРНК (mRNA)] переносит генетическую информацию из клеточного ядра в цитоплазму. Ее транскрипты также сильно модифицируются в ядре (созревание мPHK, см. с. 242) Так как мРНК считывается на рибосоме кодон за кодоном она не должна складываться в стабильную третичную структуру. Спариванию оснований препятствуют белки, ассоциированные с мРНК. Из-за различного объема информации, которую могут нести мРНК, РНК этого типа сильно варьируют по размерам. Для мРНК характерно короткое время жизни, так как они быстро распадаются после трансляции. В сплайсинге предшественников мРНК принимают участие малые ядерные РНК [мяРНК (snRNA, от англ. small nuclear RNA)]. Они ассоциированы c рядом белков, образуя «сплайсомы».
Б. Транспортные РНК ( tRNAPhe )
Транспортные РНК [тРНК (tRNA)] участвуют в процессе трансляции в качестве промежуточного связующего звена между нуклеиновыми кислотами и белками. Это небольшие молекулы РНК из 70-90 нуклеотидов, которые с помощью своих антикодонов "узнают" за счет спаривания оснований определенные кодоны на мРНК. На 3'-конце (ССА-3') они несут ту аминокислоту, которая согласно генетическому коду соответствует очередному кодону мРНК.
Последовательность оснований и третичная структура фенилаланинспецифичной тРНК (tRNAPhe) из дрожжей являются типичными для всех тРНК. В молекуле этой тРНК содержится довольно много минорных и модифицированных оснований (1, выделены темно-зеленым цветом). К ним относятся псевдоуридин (ψ), дигидроуридин (D), тимидин (T), встречающийся обычно в ДНК, а также множество метилированных нуклеотидов, таких, например, как 7-метилгуанидин (m7G) и входящий в состав антикодона 2'-О-метилгуанидин (m2G). Конформацию молекулы стабилизируют многочисленные пары оснований, часть из которых не соответствуют общим принципам спаривания оснований (2) (неканонические пары).