Глава 14. Биосинтез белка
Одной из глобальных задач современной биологии и ее новейших разделов: молекулярной биологии, биоорганической химии, физико-химической биологии – является выяснение молекулярных основ и тонких механизмов синтеза белка, содержащего сотни, а иногда и тысячи остатков L-амино-кислот. Последние располагаются, как это установлено, не хаотично, а в строго заданной последовательности, обеспечивая тем самым уникальность структуры синтезированной белковой молекулы, наделенной уникальной функцией. Другими словами, механизм синтеза должен обладать весьма тонкой и точной кодирующей системой, которая автоматически программирует включение каждого аминокислотного остатка в определенное место полипептидной цепи. Установлено, что кодирующая система однозначно определяет первичную структуру, в то время как вторичная и третичная структуры белковой молекулы определяются физико-химическими свойствами и химической структурой радикалов аминокислот в полипептиде.
Первоначально представляли, что синтез белка могут катализировать те же протеолитические ферменты, которые вызывают и его гидролиз, но путем обратимости химической реакции. Однако оказалось, что синтетические и катаболические реакции протекают не только различными путями, но даже в разных субклеточных фракциях. Не подтвердилась также гипотеза о предварительном синтезе коротких пептидов с последующим их объединением в одну полипептидную цепь. Более правильным оказалось предположение, что для синтеза белка требуются источники энергии, наличие активированных свободных аминокислот и нескольких типов клеточных нуклеиновых кислот.
В выяснение молекулярных механизмов синтеза белка определенный вклад внесли российские биохимики. Так, в лаборатории А.Е. Браунштейна было впервые указано на участие АТФ в синтезе квазипептидных связей (на примерах гиппуровой кислоты, глутамина, глутатиона и ацетанилида). В.Н. Орехович еще в 50-е годы установил, что перенос аминоацильных или пептидильных группировок на NH2-группу аминокислот может осуществляться не только с амидной или пептидной, но и со сложноэфирной связи. Как будет показано далее, именно этот механизм лежит в основе реакции транспептидирования в 50S рибосоме в стадии элонгации синтеза белка.
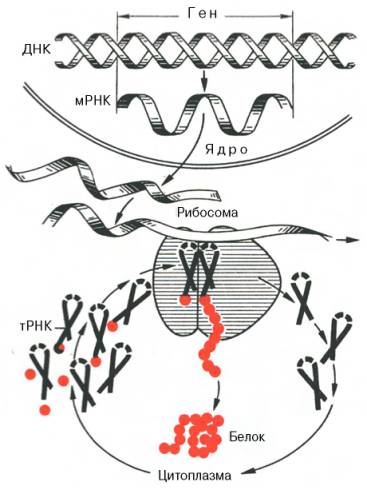
Рис. 14.1. Принципиальная схема биосинтеза белка (по А.С. Спирину).
Красные кружочки - свободные аминокислоты и их остатки в составе полипептидной цепи.
Значительно позже были получены доказательства, что в синтезе белка, протекающем в основном в цитоплазме, решающую роль играют нуклеиновые кислоты, в частности ДНК. После того как было установлено, что ДНК является носителем и хранителем наследственной информации, был поставлен вопрос о том, каким образом эта генетическая информация, записанная (зашифрованная) в химической структуре ДНК, трансформируется в фенотипические признаки и функциональные свойства живых организмов, передающиеся по наследству. В настоящее время можно дать однозначный ответ на этот вопрос: генетическая информация программирует синтез специфических белков, определяющих в свою очередь специфичность структуры и функции клеток, органов и целостного организма (рис. 14.1). В природе, как известно, существует два типа биополимерных макромолекул: так называемые неинформативные биополимеры (они представлены повторяющимися мономерными единицами и/или разветвленными структурами, например полисахариды, поли-АДФ-рибоза, пеп-тидогликаны, гликопротеины) и информативные биополимеры, несущие первичную генетическую информацию (нуклеиновые кислоты) и вторичную генетическую, точнее фенотипическую, информацию (белки). Эти общие представления могут быть выражены следующей последовательностью событий (поток информации):
ДНК –> РНК –> Белок –> Клетка –> Организм
Значительный вклад в современные представления о месте, факторах и механизме синтеза белка внесли исследования Т. Касперсона, М. Хоглан-да, П. Берга, П. Замечника, С. Очоа, М. Ниренберга, Н. Горовица, Ф. Гауровица, С. Вейсса и российских биохимиков А.А. Баева, А.Н. Белозерского, А.С. Спирина и др.
Не останавливаясь на всех исторических аспектах развития этой важнейшей проблемы, следует напомнить, что еще в 40-х годах было установлено, что ДНК локализована в ядре клетки, в то время как синтез белка протекает главным образом в микросомах цитоплазмы. Первые экспериментальные доказательства необходимости нуклеиновых кислот для синтеза белка были получены в лаборатории Т. Касперсона. Было показано также, что присутствующие в цитоплазме рибонуклеиновые кислоты контролируют синтез цитоплазматических белков. Таким образом, уже тогда вырисовывалась картина тесной связи между ДНК, локализованной в ядре , и синтезом белка, протекающим в цитоплазме и регулирующимся рибонуклеиновыми кислотами, которые были открыты как в цитоплазме, так и в ядре. На основании этих чисто морфологических данных было сделано заключение, полностью подтвержденное в настоящее время, что биосинтез белка, хотя непосредственно и регулируется рибонуклеиновыми кислотами, опосредованно связан с контролирующим влиянием ДНК ядра и что РНК сначала синтезируется в ядре, затем поступает в цитоплазму, где выполняет роль матрицы в синтезе белка. Полученные значительно позже экспериментальные данные подтвердили гипотезу о том, что основными функциями нуклеиновых кислот являются хранение генетической информации и реализация этой информации путем программированного синтеза специфических белков.
В последовательности ДНК —> РНК —> Белок недоставало сведений о том, каким образом происходят расшифровка наследственной информации и синтез специфических белков, определяющих широкое разнообразие признаков живых существ. В настоящее время выяснены основные процессы, посредством которых осуществляется передача наследственной информации: репликация, т.е. синтез ДНК на матрице ДНК; транскрипция, т.е. синтез РНК на матрице ДНК или перевод языка и типа строения ДНК на молекулу РНК (см. ранее), и трансляция – процесс, в котором генетическая информация, содержащаяся в молекуле мРНК, направляет синтез соответствующей аминокислотной последовательности в белке. Напомним, однако, что многие тонкие механизмы транскрипции и трансляции окончательно еще неясны.