Биосинтез пуринов и пиримидинов
Основания, содержащиеся в нуклеиновых кислотах, являются производными ароматических гетероциклических соединений пурина и пиримидина. Путь биосинтеза нуклеиновых оснований довольно сложен, однако этот процесс жизненно необходим почти для всех клеток. Сборка нуклеиновых оснований представлена здесь схематически. Полная схема реакций приведена на сс. 405 и 406.
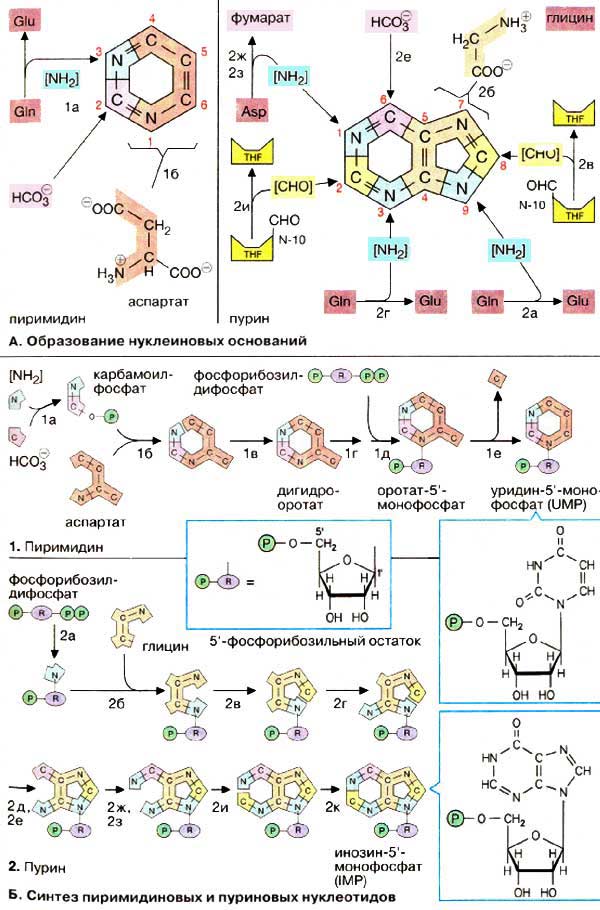
А. Образование нуклеиновых оснований
Пиримидиновое кольцо собирается из трех компонентов: атомы азота N-1 и углерода с С-4 по С-6 поставляются аспартатом. С-2 происходит из НСО3-, а второй атом азота (N-3) — из амидной группы глутамина.
Синтез пуринового кольца идет сложнее: единственным крупным предшественником является глицин, из которого происходят С-4 и С-5, а также N-7. Все другие атомы кольца поставляются отдельно: С-6 происходит из НСО3-, амидная группа глутамина дает атомы N-3 и N -9, донором аминогруппы для N-1 выступает аспартат, переход как и в цикле мочевины, в фумарат. Наконец, атомы углерода С-2 и С-8 происходят из формильной группы N10-формилтетрагидрофолата.
Б. Биосинтез пиримидиновых и пуриновых нуклеотидов
Центральными промежуточными продуктами биосинтеза предшественников нуклеиновых кислот являются мононуклеотид уридинмонофосат [УМФ (UMP)] для пиримидинового ряда и инозинмонофосфат [ИМФ (IMP), основание: гипоксантин) для пуринов. Путь синтеза различен для пиримидиновых и пуриновых оснований. В первом случае строится прежде всего пиримидиновое кольцо и затем к нуклеотиду присоединяется рибозо-5'-фосфат. Синтез пуриновых нуклеотидов, напротив, начинается с рибозо-5'-фосфата и исходя из него шаг за шагом формируется кольцо.
Непосредственными предшественниками в синтезе пиримидинового кольца являются карбамоилфосфат, который образуется из глутамина и НСО3- (1а), и аспартат. После образования N-карбамоиласпартата (1б) происходит замыкание кольца с образованием дигидрооротата (1в). У млекопитающих стадии от 1а до 1в проходят в цитоплазме и катализируются одним полифункциональным ферментом. На следующей стадии (1г) дигидрооротат окисляется флавинмо-нонуклеотидзависимой дегидрогеназой в оротат, который связывается с фосфорибозилдифосфатом (PRPP) с образованием нуклеотида оротидин-5'-монофосфата [ОМФ (ОМР)], декарбоксилирование которого приводит к уридин-5'-монофосфату [УМФ (UMP)].
Пуриновый биосинтез начинается с фосфорибозилдифосфата (названия отдельных промежуточных продуктов перечислены на с. 406). Сначала присоединяется аминогруппа, которая впоследствии в кольце становится N-9 (2а). Глицин и формильная группа N10-формилтетрагидрофолата поставляют недостающие атомы пятичленного кольца (2б, 2в). Прежде чем это кольцо замкнется (2е), присоединяются атомы N-3 и N-6 шестичленного кольца (2г, 2д). Затем построение кольца продолжается путем присоединения N-1 и С-2. На последней стадии шестичленное кольцо замыкается с образованием инозин-5'-монофосфата [ИМФ (IMP)], который, однако, не накапливается, а быстро превращается в АМФ и ГМФ. Эти реакции и синтез других нуклеотидов рассмотрены на с. 192.
Дополнительная информация
Механизм регуляции бактериальной аспартат-карбамоилтрансферазы с участием АТФ и ЦТФ изучен достаточно подробно. В организме животных ключевым ферментом пиримидинового синтеза является не аспартат-карбамоилтрансфераза, а карбамоилфосфатсинтаза. Она активируется АТФ и фосфорибозилдифосфатом (PRPP) и тормозится УТФ. Регуляция синтеза пуринов также основана на ингибировании конечным продуктом: образование PRPP из рибозо-5'-фосфата тормозится АДФ и ГДФ. Аналогичным образом АМФ и ГМФ тормозят стадию 2а.