Регуляция углеводного обмена
А. Регуляция углеводного обмена
У высших организмов обмен углеводов подвержен сложным механизмам регуляции, в которых участвуют гормоны, метаболиты и коферменты. Представленная здесь схема относится к печени, которая занимает в углеводном метаболизме центральное место (см. с. 302). Некоторые из представленных механизмов не действуют в других тканях.
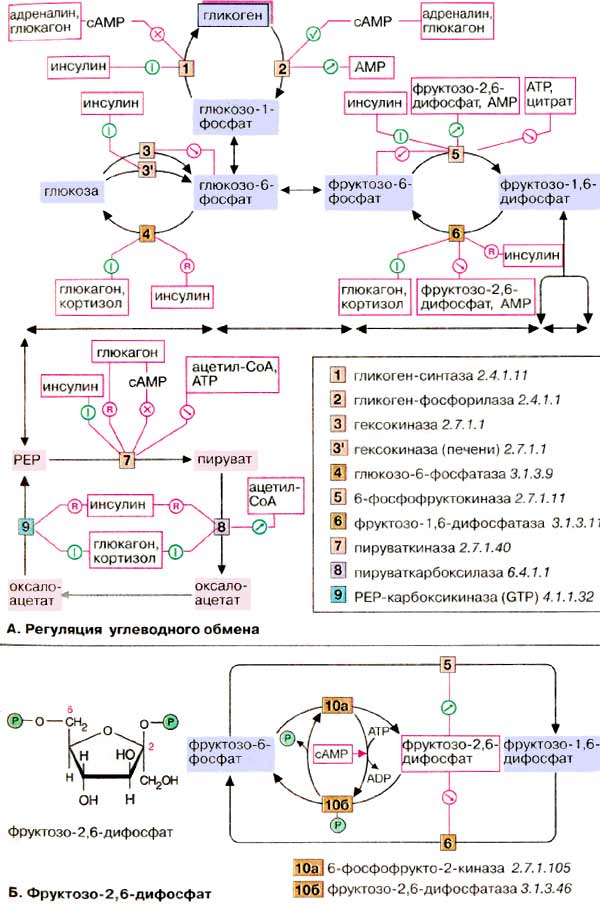
Одной из важнейших функций клеток печени является накопление избыточной глюкозы в виде гликогена и ее быстрое высвобождение по мере метаболической необходимости (буферная функция). После полной мобилизации запасов гликогена печень может поставлять глюкозу за счет синтеза de novo (глюконеогенез, см. сс. 156, 232). Кроме того, как и все ткани, она потребляет глюкозу путем гликолиза. Функции накопления (синтеза) глюкозы в виде гликогена и его распада должны быть взаимосогласованы. Таким образом, совершенно невозможно одновременное протекание гликолиза и глюконеогенеза, как и синтеза и деградации гликогена. Согласование процессов обеспечивается тем, что синтез (анаболизм) и распад (катаболизм) катализируются двумя различными ферментами и контролируются независимо. На схеме показаны только эти ключевые ферменты.
Гормоны. К гормонам, которые влияют на углеводный обмен, принадлежат пептиды инсулин и глюкагон, глюкокортикоид кортизол и катехоламин адреналин. Инсулин индуцирует (см. с. 120) синтез de novo гликоген-синтазы [1], а также некоторых ферментов гликолиза [3, 5, 7]. Одновременно инсулин подавляет синтез ключевых ферментов глюконеогенеза (репрессия, [4, 6, 8, 9]). Глюкагон как антагонист инсулина действует в противоположном направлении: индуцирует ферменты глюконеогенеза [4, 6, 8, 9] и репрессирует пируваткиназу [7], ключевой фермент гликолиза. Другие аффекты глюкагона основаны на взаимопревращении ферментов и опосредованы вторичным мессенджером цАМФ (сАМР, см. с. 114). По этому механизму тормозится синтез гликогена [1] и активируется расщепление гликогена [2]. Подобным образом действует и адреналин. Торможение пируваткиназы [7] глюкагоном также обусловлено взаимопревращением ферментов.
Глюкокортикоиды, прежде всего кортизол, индуцируют все ключевые ферменты глюконеогенеза [4, 6, 8, 9]. Одновременно они индуцируют ферменты деградации аминокислот и обеспечивают тем самым глюконеогенез исходными соединениями.
Метаболиты. Высокие концентрации АТФ (АТР) и цитрата тормозят гликолиз путем аллостерической регуляции фосфофруктокиназы. Кроме того, АТФ тормозит пируваткиназу. Ингибитором пируваткиназы является ацетил-КоА. Все эти метаболиты образуются при распаде глюкозы (торможение конечным продуктом). АМФ (AMP), сигнал дефицита АТФ, активирует расщепление гликогена и тормозит глюконеогенез.
Б. Фруктозо-2,6-дифосфат
Важную роль в обмене веществ в печени играет фруктозо-2,6-дифосфат. Это сигнальное вещество образуется в незначительных количествах из фруктозо-6-фосфата и выполняет чисто регуляторную функцию: стимулирует гликолиз путем активации фосфофруктокиназы и подавляет глюконеогенез с помощью торможения фруктозо-1,5-дифосфатазы.
Образование и распад фруктозо-2,6-дифосфата катализируются одним и тем же белком [10а и б]. В нефосфорилированной форме этот белок вызывает образование фруктозо-2,6-дифосфата [10а]. После фосфорилирования цАМФ-зависимой киназой он действует как фосфатаза [10б] и катализирует превращение фруктозо-2,6-дифосфата в фруктозо-6-фосфат. В присутствии адреналина и глюкагона в клетках печени повышается уровень цАМФ, т.е. оба гормона воздействуют как на гликолиз, так и на глюконеогенез. Суммарным результатом является быстрое повышение уровня глюкозы в крови.