Механизмы регуляции метаболических процессов
А. Основные механизмы регуляции метаболических процессов
Активность всех путей обмена веществ постоянно регулируется, что обеспечивает соответствие синтеза и деградации метаболитов физиологическим потребностям организма. В этом разделе рассматриваются механизмы такой регуляции. Более детально вопросы регуляции клеточного метаболизма представлены на сс. 118-123.
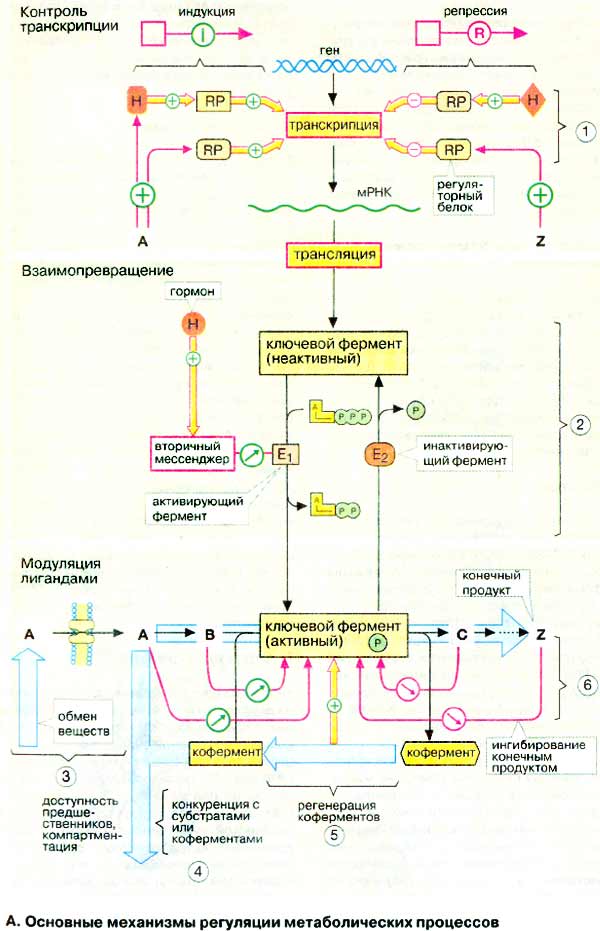
Поток метаболитов в обмене веществ определяется прежде всего активностью ферментов. Для воздействия на тот или иной путь достаточно регулировать активность фермента, катализирующего наиболее медленную стадию. Такие ферменты, называемые ключевыми ферментами, имеются в большинстве метаболических путей. Активность ключевого фермента регулируется на трех независимых уровнях,
Взаимопревращение. Значительно быстрее, чем контроль транскрипции, действует взаимопревращение ключевых ферментов (2). В этом случае фермент присутствует в клетке в неактивной форме. При метаболической потребности по сигналу извне и при посредничестве вторичного мессенджера активирующий фермент (E1) переводит ключевой фермент в каталитически активную форму. Если потребность в этом пути обмена веществ отпадает, инактивирующий фермент (E2) снова переводит ключевой фермент в неактивную форму. Процесс взаимопревращения в большинстве случаев состоит в АТФ-зависимом фосфорилировании ферментных белков протеинкиназой и соответственно дефосфорилировании фосфатазой. В большинстве случаев более активна фосфорилированная форма фермента, однако встречаются также и противоположные случаи.
Модуляция лигандами. Важным параметром, контролирующим протекание метаболического пути, является потребность в первом реагенте (здесь это метаболит А). Доступность метаболита А возрастает с повышением активности метаболического пути (3), в котором образуется А, и падает с повышением активности других путей (4), в которых А расходуется. Доступность А может быть ограничена в связи с его транспортом в другие отделы клетки.
Часто лимитирующим фактором является также доступность кофермента (5). Если кофермент регенерируется по второму независимому пути, этот путь может лимитировать скорость основной реакции. Таким образом, например, гликолиз и цитратный цикл регулируются доступностью НАД+. Так как НАД+ регенерируется в дыхательной цепи, последняя регулирует катаболизм глюкозы и жирных кислот (контроль дыхания, см. с. 146).
Наконец, активность ключевого фермента может регулироваться лигандом (субстратом, конечным продуктом реакции, коферментом, другим эффектором) как аллостерическим эффектором путем связывания его не в самом активном центре, а в другом месте фермента, и вследствие этого изменением ферментативной активности (6, см. с. 118). Ингибирование ключевого фермента часто вызывается конечными продуктами реакции соответствующей метаболической цепи (ингибирование по типу обратной связи) или метаболитом, участвующим в другом пути. Стимулировать активацию фермента может также первый реагент реакционной цепи.