Минорные нуклеозиды
МИНОРНЫЕ НУКЛЕОЗИДЫ, пуриновые и пиримидино-выс нуклеозиды, входящие в молекулы прир. нуклеиновых к-т в относительно небольших кол-вах. В отличие от наиб. распространенных нуклеозидов (аденозина, гуанозина, ури-дина, тимидина и цитидина) содержат модифицир. гетеро-циклич. основания и(или) остатки рибозы или дезоксири-бозы.
Минорные нуклеозиды обнаружены практически во всех нуклеиновых к-тах. Наиб. высокое содержание минорных нуклеозидов наблюдается у эукариотич. транспортных РНК (тРНК), у к-рых доля минорных нуклеозидов достигает 20-25% от общего кол-ва нуклеозидов. Значительно меньше (1-2%) минорных нуклеозидов в рибосомных РНК (рРНК). У последних минорные нуклеозиды сосредоточены в огранич. числе мест. В отличие от РНК содержание минорных нуклеозидов в ДНК разных организмов сильно варьирует. Так, в ДНК насекомых минорные нуклеозиды достоверно не обнаружены, в ДНК позвоночных их содержится 1-2%, а у растений эта величина достигает 3-8%. Минорные нуклеозиды находятся в ДНК обычно не в уникальных, а в повторяющихся последовательностях.
Характер модификаций в минорных нуклеозидах варьирует очень широко, но в значит. степени преобладает N-, О- и С-метилирование оснований.
В ДНК растит. и животных тканей встречается, как правило, только 5-мстил-2'-дезоксицитидин (обозначается m5dC; ф-ла I, X = Н, Y = СН3; dRib-остаток 2'-дезокси-рибозы), а у прокариот (бактерии и синезеленые водоросли)-также его изомер (m4dC; ф-ла I, X = СН3, Y = H) и N6-метил-2'-дезоксиаденозин (m6dA; ф-ла II); в меньших кол-вах присутствуют N1-метил- и N2 -диметил-2'-дезоксигуанозин (m1dG и m22dG; ф-ла III, соотв. X = СН3, Y = Н и X = Н, Y = СН3). Единственная отличная от метилирования модификация в ДНК состоит в замене 5-метильной группы тимина в ДНК Т-четных фагов (вирусов бактерий) на гидроксиметильную группу, к-рая м. б. также гликозилирована.
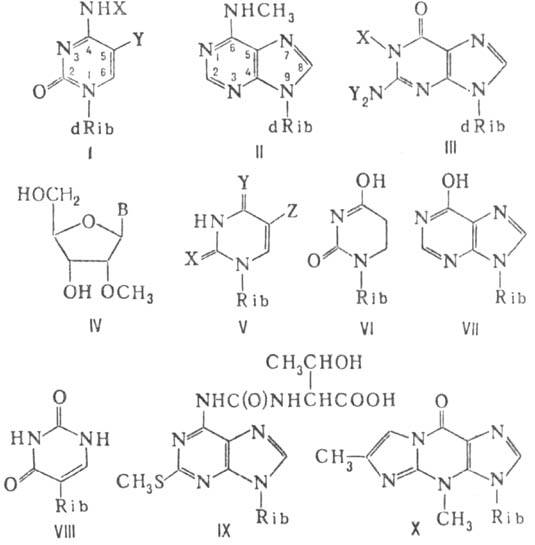
В РНК бактерий метилированию подвергаются в осн. гетероциклич. основания, тогда как у высших организмов минорные нуклеозиды в РНК представлены гл. обр. 2-О-метилрибозидами (Nm; ф-ла IV, В-основание). Наиб. структурное разнообразие минорных нуклеозидов характерно для тРНК, где м. б. модифицированы все четыре типа нуклеозидных звеньев. При этом минорные нуклеозиды образуются не только в результате присоединения к ге-тероциклу или к экзоциклич. аминогруппе разл. группировок, в простейшем случае - группы СН3 (напр., 5-мети-луридин-т5U; ф-ла V, X = Y = О, Z = СН3, Rib-остаток рибозы), но также в результате восстановления двойной связи в положении 5,6 пиримидина (5,6-дигидроуридин-hU; ф-ла VI).
Кроме того, минорные нуклеозиды могут образовываться в результате замены карбонильного атома кислорода на атом серы (2-или 4-тиоуридин-S2U и S4U; ф-ла V, соотв. X = S, Y = О, Z = Н и X = О, Y = S, Z = H) или аминогруппы на гидро-ксил (инозин-ф-ла VII), а также в результате изменения взаимного расположения урацильного и углеводного остатков с образованием С-гликозидной связи (5-рибозилурацил, или псевдоуридин,-y; ф-ла VIII).
Известно немало минорных нуклеозидов, модифицированных по двум позициям (напр., 5-мстил-2-тиоуридин-m5s2U; ф-ла V, X = S, Y = О, Z = СН3), а также минорные нуклеозиды с заместителями, образующимися в результате ряда последовательных модификаций, напр. производное уридина (mcm5s2U; ф-ла V, X = S, Y = О, Z = CH2COOCH3) и аденозина (ms2t6A; ф-ла IX), в т.ч. циклизации (напр., виозин-W; ф-ла X). Такие гипермодифицированные минорные нуклеозиды всегда занимают в молекуле тРНК позицию 37, т.е. соседствуют с 3'-концевым нуклеозидом антикодона- участка молекулы тРНК, состоящего из трех нуклсотидов и узнающего соответствующий ему участок из трех нуклеотидов (кодон) в молекуле матричной РНК при трансляции (синтезе белка на РНК-матрице).
Образование минорных нуклеозидов в РНК и ДНК происходит на поли-нуклеотидном уровне (т. е. после того, как полимерная цепь уже сформирована) путем переноса соответствующих групп спeциализир. ферментами с доноров в определенные положения нуклеиновой к-ты. Полнее всего изучено метилиро-вание, к-рое во всех случаях (за одним исключением) осуществляется с участием S-аденозилметионина - универсального донора метильной группы. Ферменты, катализирующие образование минорных нуклеозидов, высокоспецифичны к определенному нуклеозиду и к разным позициям его в молекуле.
Описано влияние ДНК с метилир. основаниями на ряд процессов (экспрессия генов, репликация и др.), однако мол. механизмы, лежащие в основе этого явления, не выяснены. Несколько больше известно о роли минорных нуклеозидов в ф-ции тРНК. Возрастание кол-ва минорных нуклеозидов в тРНК по мере усложнения организмов указывает на то, что они имеют отношение к добавочным, напр. регуляторным, ф-циям тРНК, поскольку фундам. механизмы переноса аминокислот сходны во всех организмах. Даже самая простая модификация, напр. ме-тилирование, влияет на конформацию молекулы, меняя способность к образованию водородных связей и стэкинг-взаимод. между основаниями. Т. обр., влияние минорных нуклеозидов на ф-ции тРНК осуществляется, вероятно, через изменение ее конформации, причем разные по структуре и положению минорные нуклеозиды, безусловно, играют разл. роль. Напр., неизменность локализации гипермодифицированных минорных нуклеозидов с 3'-стороны от антикодона позволяет предположить их участие в считывании кодона. Минорные нуклеозиды в рРНК, по-видимому, участвуют в приобретении молекулой конформации, необходимой для взаимод. с рибосомными белками.
Исп. литература для статьи «МИНОРНЫЕ НУКЛЕОЗИДЫ»: Органическая химия нуклеиновых кислот, М., 1970; Романов А. Г., Ванюшин Б. Ф., "Биологические науки", 1980, № 11, с. 5-20; их же, там же, 1981, № 1, с. 5 13; Saenger W., Principles of nucleic acid structure, N. Y., 1984.
Т. В. Вeнкстерн.