Са2+-мессенджерная система
Ионам Са2+ принадлежит центральная роль в регуляции многих клеточных функций. Изменение концентрации внутриклеточного свободного Са2+ является сигналом для активации или ингибирования ферментов, которые в свою очередь регулируют метаболизм, сократительную и секреторную активность, адгезию и клеточный рост. Источники Са2+ могут быть внутри- и внеклеточными. В норме концентрация Са2+ в цитозоле не превышает 10–7 М, и основными источниками его являются эндоплазмати-ческий ретикулум и митохондрии. Нейрогормональные сигналы приводят к резкому повышению концентрации Са2+ (до 10–6 М), поступающего как извне через плазматическую мембрану (точнее, через потенциалзависимые и рецепторзависимые кальциевые каналы), так и из внутриклеточных источников. Одним из важнейших механизмов проведения гормонального сигнала в кальций–мессенджерной системе является запуск клеточных реакций (ответов) путем активирования специфической Са2+-кальмодулин-зависимой протеинкиназы. Регуляторной субъединицей этого фермента оказался Са2+-связывающий белок кальмодулин (мол. масса 17000). При повышении концентрации Са2+ в клетке в ответ на поступающие сигналы специфическая протеинкиназа катализирует фосфорилирование множества внутриклеточных ферментов – мишеней, регулируя тем самым их активность. Показано, что в состав киназы фосфорилазы b, активируемой ионами Са2+, как и NO-синтазы, входит кальмодулин в качестве субъединицы. Кальмодулин является частью множества других Са2+-свя-зывающих белков. При повышении концентрации кальция связывание Са2+ с кальмодулином сопровождается конформационными его изменениями, и в этой Са2+-связанной форме кальмодулин модулирует активность множества внутриклеточных белков (отсюда его название).
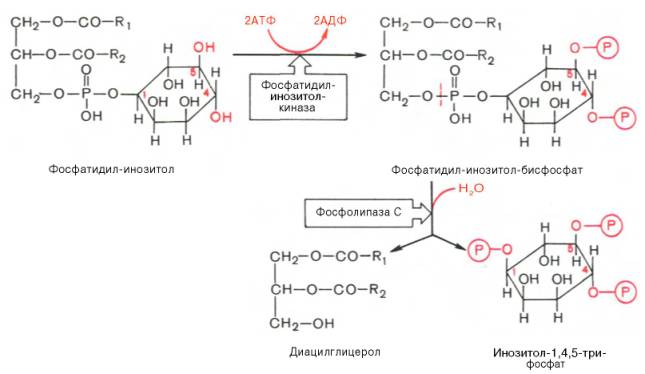
К внутриклеточной системе мессенджеров относят также производные фосфолипидов мембран эукариотических клеток, в частности фосфорили-рованные производные фосфатидилинозитола. Эти производные освобождаются в ответ на гормональный сигнал (например, от вазопрессина или тиротропина) под действием специфической мембраносвязанной фосфо-липазы С. В результате последовательных реакций образуются два потенциальных вторичных мессенджера – диацилглицерол и инозитол-1,4,5-три-фосфат.
Биологические эффекты этих вторичных мессенджеров реализуются по-разному. Действие диацилглицерола, как и свободных ионов Са2+, опосредовано через мембраносвязанный Са-зависимый фермент протеин-киназу С, которая катализирует фосфорилирование внутриклеточных ферментов, изменяя их активность. Инозитол-1,4,5-трифосфат связывается со специфическим рецептором на эндоплазматическом ретикулуме, способствуя выходу из него ионов Са2+ в цитозоль.
Таким образом, представленные данные о вторичных мессенджерах свидетельствуют о том, что каждой из этих систем посредников гормонального эффекта соответствует определенный класс протеинкиназ, хотя нельзя исключить возможности существования тесной связи между этими системами. Активность протеинкиназ типа А регулируется цАМФ, протеинкиназы G – цГМФ; Са2+-кальмодулинзависимые протеинкиназы находятся под контролем внутриклеточной [Са2+], а протеинкиназа типа С регулируется диацилглицеролом в синергизме со свободным Са2+ и кислыми фосфолипидами. Повышение уровня какого-либо вторичного мес-сенджера приводит к активации соответствующего класса протеинкиназ и последующему фосфорилированию их белковых субстратов. В результате меняется не только активность, но и регуляторные и каталитические свойства многих ферментных систем клетки: ионных каналов, внутриклеточных структурных элементов и генетического аппарата.
Известно, что эффект стероидных гормонов реализуется через генетический аппарат путем изменения экспрессии генов. Гормон после доставки с белками крови в клетку проникает (путем диффузии) через плазматическую мембрану и далее через ядерную мембрану и связывается с внутриядерным рецептором–белком. Комплекс стероид–белок затем связывается с регуляторной областью ДНК, с так называемыми гормончувствительны-ми элементами, способствуя транскрипции соответствующих структурных генов, индукции синтеза белка de novo (см. главу 14) и изменению метаболизма клетки в ответ на гормональный сигнал.
Следует подчеркнуть, что главной и отличительной особенностью молекулярных механизмов действия двух основных классов гормонов является то, что действие пептидных гормонов реализуется в основном путем посттрансляционных (постсинтетических) модификаций белков в клетках, в то время как стероидные гормоны (а также тиреоидные гормоны, ретиноиды, витамин D3-гормоны) выступают в качестве регуляторов экспрессии генов. Это обобщение, однако, не является абсолютным, и здесь возможны модификации, рассмотренные при описании отдельных гормонов.