Метаболизм пептидных гормонов А. Биосинтез
В отличие от стероидов пептидные и белковые гормоны являются первичными продуктами биосинтеза. Соответствующая информация считывается с ДНК (DNA) на стадии транскрипции, а синтезированная гяРНК (hnRNA) освобождается от интронов за счет сплайсинга (1). мРНК (mRNA) кодирует последовательность пептида, который чаще всего существенно превышает по молекулярной массе зрелый гормон. Исходная аминокислотная цепь включает сигнальный пептид и пропептид — предшественник гормона. Трансляция мРНК происходит на рибосомах по обычной схеме (2). Вначале синтезируется сигнальный пептид. Его функция состоит в том, чтобы связать рибосомы на шероховатом эндоплазматическом ретикулуме [ШЭР (rER)] и направить растущую пептидную цепь в просвет ШЭР (3). Синтезированный продукт является предшественником гормона, прогормоном. Созревание гормона происходит путем ограниченного протеолиза и последующей (посттрансляционной) модификации, например образования дисульфидных мостиков, гликозилирования и фосфорилирования (см. с. 226) (4). Зрелый гормон депонируется в клеточных везикулах, откуда секретируется по мере необходимости за счет экзоцитоза.
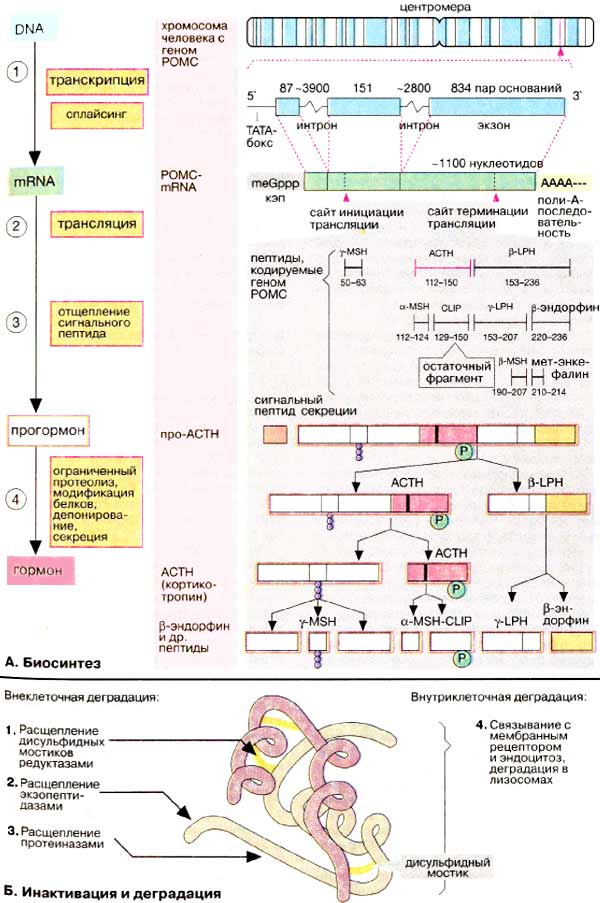
Биосинтез пептидных и белковых гормонов и их секреция находятся под контролем иерархической системы гормональной регуляции. В этой системе в качестве вторичного мессенджера принимают участие ионы кальция; увеличение концентрации кальция стимулирует синтез и секрецию гормонов.
Анализ гормональных генов показывает, что иногда многие совершенно разные пептиды и белки кодируются одним и тем же геном. Одним из наиболее изученных является ген проопиомеланокортина [ПОМК (POMC)]. Наряду с нуклеотидной последовательностью, соответствующей кортикотропину [адренокортикотропный гормон, АКТГ (АСТН)], этот ген включает перекрывающиеся последовательности, кодирующие ряд небольших пептидных гормонов, а именно α-, β- и γ-меланотропинов [МСГ (MSH)], β- и γ- липотропинов (ЛПГ (LPH)], β-эндорфина и мет-энкефалина. Последний гормон может также образовываться из β-эндорфина. Прогормоном для этого семейства является так называемый полипротеин. Сигнал о том, какой пептид должен быть получен и секретирован, поступает из системы регуляции после завершения синтеза препропептида. Наиболее важным секретируемым продуктом, полученным из гипофизарного полипротеина кодируемого геном ПОМК, является гормон кортикотропин (АКТГ), стимулирующий секрецию кортизола корой надпочечников. Биологические функции других пептидов до конца не выяснены.
Б. Инактивация и деградация
Деградация пептидных гормонов часто начинается уже в крови или на стенках кровеносных сосудов, особенно интенсивно этот процесс идет в почках. Некоторые пептиды, содержащие дисульфидные мостики, например инсулин, могут инактивироваться за счет восстановления остатков цистина (1), Другие белково-пептидные гормоны гидролизуются протеиназами, а именно экзо- (2) (по концам цепи) и эндопептидазами (3). Протеолиз приводит к образованию множества фрагментов, некоторые из которых могут проявлять биологическую активность. Многие белково-пептидные гормоны удаляются из системы циркуляции за счет связывания с мембранным рецептором и последующего эндоцитоза гормон-рецепторного комплекса. Деградация таких комплексов происходит в лизосомах, конечным продуктом деградации являются аминокислоты, которые вновь используются в качестве субстратов в анаболических и катаболических процессах.
Дополнительная информация
Липофильные и гидрофильные гормоны имеют различный полупериод существования в системе циркуляции (точнее биохимический полупериод, t1/2). По сравнению с гидрофильными гормонами (t1/2 несколько минут или часов) липофильные гормоны живут существенно дольше (t1/2 составляет несколько часов или дней). Биохимический полупериод гормонов зависит от активности системы деградации. Воздействие на систему деградации лекарственными препаратами или повреждение тканей может вызвать изменение скорости распада, а следовательно, и концентрации гормонов.