Пентозофосфатный цикл
ПЕНТОЗОФОСФАТНЫЙ ЦИКЛ (пентозный путь, гексо-зомонофосфатный шунт, фосфоглюконатный путь), совокупность обратимых ферментативных р-ций, в результате к-рых происходит окисление глюкозы до CO2 с образованием восстановленного никотинамидадениндинуклеотид-фосфата (НАДФН) и H + , а также синтез фосфорилир. Сахаров, содержащих от 3 до 7 атомов С.
Пентозофосфатный цикл осуществляется в цитозоле (жидкой фазе) клеток животных, растений (особенно в темноте) и микроорганизмов. У растений часть р-ций пентозофосфатного цикла участвует также в образовании гексоз при фотосинтезе.
РЕАКЦИИ ПЕНТОЗОФОСФАТНОГО ЦИКЛА И ФЕРМЕНТЫ ИХ КАТАЛИЗИРУЮЩИЕ
| Номер р-ции на схеме | Р-ции | Фермент, катализирующий р-цию | ||
| 1 | 6 Глюкозо-6-фосфат + 6 НАДФ | Глюкозо-6-фос- фат-дегидроге-наза | ||
| 2 | 6 6-Фосфоглюконолактон | Лактоназа | ||
| 3 | 6 6-Фосфоглюконат + 6 НАДФ | 6-Фосфоглюко-нат-дегидроге-наза | ||
| 4 | 2 Рибулозо-5-фосфат | Пентозоизоме-раза | ||
| 5 | 4 Рибулозо-5-фосфат | Фосфорибулозо-эпимераза | ||
| 6 | 2 Ксилолуозо-5-фосфат + 2 Рибозо-5-фосфат | |||
| 7 | 2 Седогептулозо-7-фосфат + 2 Глицеральдегид-3-фосфат | Трансальдолаза | ||
| 8 | 2 Ксилулозо-5-фосфат + 2 Эритрозо-4-фосфат | Транскетолаза | ||
| 9 | Глицеральдегид-3-фосфат | Триозофосфат-изомераза | ||
| 10 | Дигидроксиацетонфосфат + Глицеральдегид-3-фос-фат | |||
| 11 | Фруктозе- 1, 6- дифосфат + | Фруктозо-бис-фосфатаза | ||
| 12 | 5 Фруктозо-6-фосфат | Гексозофосфат-изомераза | ||
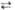 6 6-Фосфоглюко-нолактон + 6 НАДФН + 6Н+
6 6-Фосфоглюко-нолактон + 6 НАДФН + 6Н+ 6 6-Фосфоглюконат
6 6-Фосфоглюконат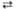 6 Риболозо-5-фосфат + 6 НАДФН + 6 H+ + 6CO2
6 Риболозо-5-фосфат + 6 НАДФН + 6 H+ + 6CO2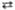 2 Рибозо-5-фосфат
2 Рибозо-5-фосфат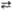 4 2-Ксилулозо-5-фосфат
4 2-Ксилулозо-5-фосфат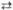 2 Седогептулозо-7-фосфат + 2 Глиперальдегид-3-фосфат
2 Седогептулозо-7-фосфат + 2 Глиперальдегид-3-фосфат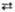 2 Эритрозо-4-фосфат + 2 Фруктозо-6-фосфат
2 Эритрозо-4-фосфат + 2 Фруктозо-6-фосфат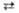 2 Глицеральдегид-3-фосфат + 2 Фруктозо-6-фосфат
2 Глицеральдегид-3-фосфат + 2 Фруктозо-6-фосфат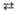 Дигидроксиацетон-фосфат
Дигидроксиацетон-фосфат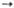 Фруктозо- 1 ,6-дифосфат
Фруктозо- 1 ,6-дифосфат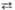 Фруктозо-6-фосфат +
Фруктозо-6-фосфат +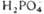
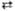 5 Глюкозо-6-фосфат
5 Глюкозо-6-фосфат Суммарный процесс: 6 Глюкозо-6-фосфат + 12 НАДФ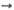 5 Глюкозо-6-фос-фат+6 CO2 + 12 НАДФН +12H+ +
5 Глюкозо-6-фос-фат+6 CO2 + 12 НАДФН +12H+ + 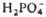
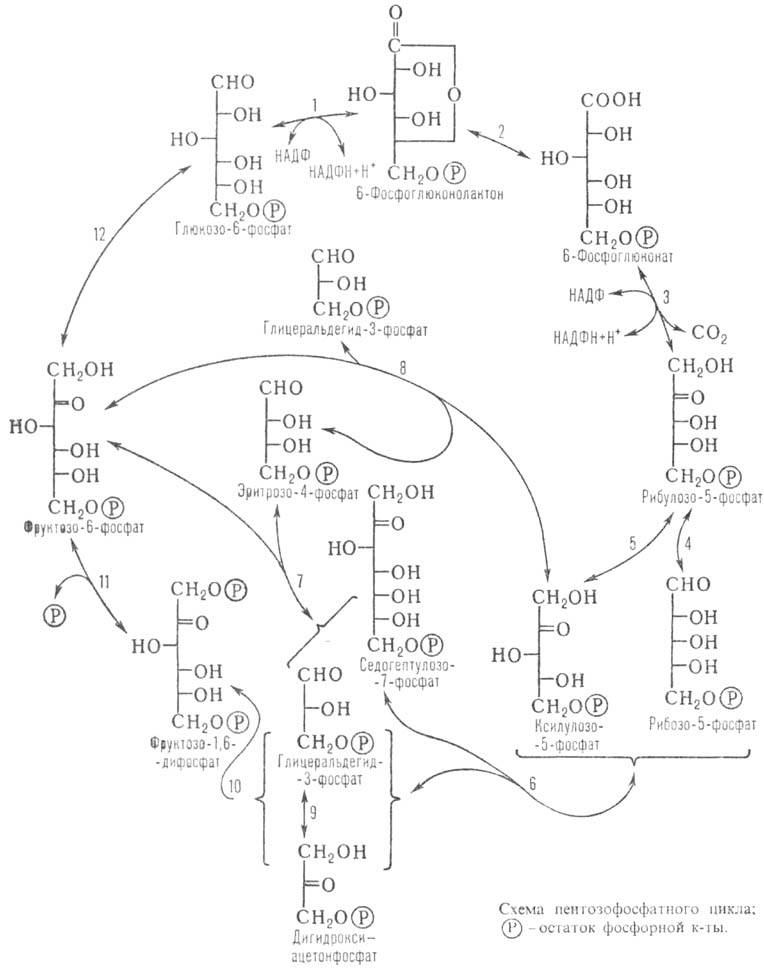
Первая (окислит.) стадия пентозофосфатного цикла (р-ции 1-3, см. схему) осуществляется с образованием НАДФН (осуществляет восстановление субстратов в организме) и рибулозо-5-фос-фата, к-рый затем превращ. в рибозо-5-фосфат (все сахара находятся в D-форме), входящий в состав молекул ряда важнейших прир. соед. (нуклеиновых к-т, нуклеотидов и др.). На неокислит. стадии пентозофосфатного цикла (остальные р-ции) в результате взаимопревращения Сахаров образуются промежут. продукты гликолиза (фруктозо-6-фосфат, глицеральдегид-3-фосфат) и таким образом осуществляется обратимая связь пентозофосфатного цикла с гликолитич. путем метаболизма глюкозы.
В отличие от др. осн. путей метаболизма углеводов (гликолиза, трикарбоновых кислот цикла)функционирование пентозофосфатного цикла нельзя представить в виде линейной последовательности р-ций, приводящей непосредственно от 1 молекулы глюкозо-6-фосфата к 6 молекулам CO2. Пентозофосфатный цикл характеризуется возможностью многообразных взаимопревращений его метаболитов, происходящих по неск. альтернативным путям. Р-ции отдельных стадий пентозофосфатного цикла (их стехиометрия) и суммарная р-ция цикла приведены в таблице.
Важная особенность пентозофосфатного цикла (в сравнении с др. путями метаболизма углеводов)-его гибкость. Если потребность в рибозо-5-фосфате значительно превышает потребность в НАДФН, то б.ч. глюкозо-6-фосфата по гликолитич. пути превращ. в глицеральдегид-3-фосфат, 1 молекула к-рого, вступая в р-ции с 2 молекулами фруктозо-6-фосфата, превращ. в 3 молекулы рибозо-5-фосфата (обращение р-ций 6-8).
В случаях, когда потребность в НАДФН и рибозо-5-фосфате сбалансирована, преобладающими становятся р-ции окислит, стадии пентозофосфатного цикла и р-ция 4. Суммарное ур-ние такого процесса:
Глюкозо-6-фосфат + 2 НАДФ + H2O 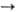 ри-бозо-5-фосфат + 2 НАДФН + 2 H+ + CO2
ри-бозо-5-фосфат + 2 НАДФН + 2 H+ + CO2
Если потребность в НАДФН значительно превышает потребность в рибозо-5-фос-фате, происходит полное окисление глю-козо-6-фосфата до CO2, включающее окислит, стадию пентозофосфатного цикла и ресинтез глюкозо-6-фос-фата из фруктозо-6-фосфата по пути глю-конеогенеза (р-ции 1-4, 6, 10-12). В этом случае суммарное ур-ние р-ции:
Глюкозо-6-фосфат + 12 НАДФ + 7 H2O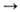 6CO2 + 12 НАДФН + 12 H+ + H3PO4
6CO2 + 12 НАДФН + 12 H+ + H3PO4
В условиях, когда потребность в НАДФН значительно превышает потребность в рибозо-5-фосфате, возможна реализация др. механизма, в соответствии с к-рым образующийся рибозо-5-фосфат превращ. не в глюкозо-6-фосфат, а в пиро-виноградную к-ту (пируват) в результате гликолиза фруктозо-6-фосфата и глице-ральдегид-3-фосфата, образующихся в р-циях 6-8. При этом образуются НАДФН, НАДН (восстановленная форма никотинамидадениндинуклеотида) и АТФ по суммарному ур-нию:
3 глюкозо-6-фосфат + 6 НАДФ + 5 НАД + + 5 H3PO4 + 8 АДФ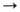 5 пируват + 3 CO2 + + НАДФН + 5 НАДН + 8 АТФ + 2 H2O + 8H +
5 пируват + 3 CO2 + + НАДФН + 5 НАДН + 8 АТФ + 2 H2O + 8H +
HАД - окисленная форма НАДН, АДФ-аденозиндифосфат Образующаяся пировиноградная к-та может далее претерпевать превращ. в цикле трикарбоновых к-т (при этом образуется АТФ) в др. р-циях в обмене в-в.
Регуляция направленности р-ций в пентозофосфатном цикле осуществляется гл. обр. ферментами, участвующими в этом цикле: избыток того или иного субстрата подавляет активность фермента, катализирующего его синтез, или активирует фермент, катализирующий его трансформацию в др. соединение.
Относит. кол-ва глюкозы, превращающиеся через пентозофосфатный цикл, неодинаковы в разных тканях. В мышцах скорость пентозофосфатного цикла очень низка, а в печени не менее 30% CO2 образуется при окислении глюкозы в пентозофосфатном цикле. В др. тканях, где активно проходит биосинтез жирных к-т и стероидов (семенниках, жировой ткани, лейкоцитах, коре надпочечников, молочной железе), доля пентозофосфатного цикла в окислит. метаболизме глюкозы также очень значительна.
Интенсивность пентозофосфатного цикла зависит от функцион. состояния ткани и от гормонального статуса (напр., в печени резко снижается при голодании из-за инактивации дегидрогеназ пентозофосфатного цикла и восстанавливается вскоре после кормления). Скорость пентозофосфатного цикла регулируется в первую очередь концентрацией НАДФН. Обе дегидрогеназы пентозофосфатного цикла (р-ции 1 и 3) чувствительны к изменению величины отношения НАДФ/НАДФН: при его величине 0,02 активность дегидрогеназ в печени максимальна, а при величине 0,01 снижается на 90%. Интенсивный пентозофосфатный цикл происходит в эритроцитах, что связано с необходимостью НАДФН-зависимого восстановления глутатиона кофактора глутатионредуктазы эритроцитов.
Нарушения функционирования нек-рых ферментов пентозофосфатного цикла приводят к развитию тяжелых заболеваний человека. Недостаточность глюкозо-6-фосфат-дегидрогеназы в эритроцитах служит причиной лек. гемолитич. анемии, а снижение активности транскетолазы в результате нарушения ее способности связывать тиамин приводит к развитию нервно-психич. расстройства синдрома Вернике Корсакова.
Открытие О. Варбугом в 1931 фермента глюкозо-6-фос-фат-дегидрогеназы, катализирующего первую р-цию пентозофосфатного цикла, сделало возможным его полную расшифровку, к-рую осуществили F Дикенс, Ф. Липман, Э. Рэкер и Б. Хорекер.
Лит Основы биохимии, пер. с англ., т. 2. M.. 1981.c 599 608; Страйер Л.. Биохимия, пер с анг.1 .т 2. M , 1985, с 95 105: Reflections on biochemistry, Oxf., 1976