Микрогетерогенность
Что такое «чистое вещество», «индивидуальное вещество», «гомогенное вещество»? По классическому определению (если отвлечься от неизбежного количества присутствия большего или меньшего количества примесей), это вещество, все молекулы которого одинаковы по структуре. Подобное определение, казалось бы, не должно вызывать ни сомнений, ни двусмысленных толкований. И так оно и есть, пока речь идет о низкомолекулярных веществах или даже о многих сравнительно высокомолекулярных. Однако уже на примере глюкозы видно, что дело тут может оказаться не таким простым.
Действительно, если речь идет о кристаллической a -D-глюкопиранозе, трудно усомниться в том, что она по всем меркам подходит под определение «индивидуальное вещество». Однако стоит только растворить эту самую глюкозу в воде, как начинается мутаротация, в результате которой мы получим раствор четырех циклических форм моносахарида. Можно ли говорить о нем, как о растворе индивидуального вещества? И да, и нет. Но, может быть, дело в том, что оно находится именно в растворе, где есть еще один компонент – растворитель?
Упарим раствор, отогнав воду в вакууме. В остатке мы получим сироп, содержащий все четыре циклические формы. Индивидуально ли такое вещество? По-видимому, нет, так как в нем присутствуют молекулы четырех структурно различных типов – четырех изомеров. При стоянии этот сироп самопроизвольно закристаллизовался, мы опять получили индивидуальную a -D-глюкопиранозу. Как же так: из смеси четырех веществ, не прибегая к химическим или физическим воздействиям, мы снова получили индивидуальное вещество? Видно, с понятием индивидуальности и в самом деле не все обстоит так просто, как кажется.
Возьмем синтетический полимер, например полиэтилен. Можно получить полиэтилен высокой степени чистоты. Индивидуален ли такой полимер? Да, в том смысле, что все его молекулы устроены одинаково: это линейные цепи из большого числа звеньев –CH 2 -CH 2 -. И нет, в том смысле, что молекулы в образце полиэтилена различаются по молекулярной массе. Эти различия могут быть велики или малы (в зависимости от молекулярно-массового распределения); но в синтетических полимерах они есть всегда, так как рост каждой отдельной цепи при их синтезе подчиняется закону случая. Таким образом, в образце полимера не только не «все молекулы одинаковы по структуре», но в нем множество различающихся по структуре молекул.
Правда, небольшие вариации молекулярной массы высокомолекулярного соединения при неизменности повторяющейся структуры – это, казалось бы, нечто сравнительно мало существенное. Трудно ведь допустить, чтобы полиэтилен с молекулярной массой 100000 сильно отличался от полиэтилена с молекулярной массой 99500 или тем более от смеси двух полиэтиленов с молекулярными массами 99500 и 100500. Так что применительно к полимерам понятие «индивидуальное вещество» приходится трактовать расширенно.
Но то синтетические полимеры. Часть биополимеров синтезируется в клетке отнюдь не по закону случая. Наиболее известный пример – белки. Сборка их полипептидных цепей происходит на рибонуклеиновой матрице, вследствие чего положение каждой аминокислоты строго детерминировано. Иначе быть не может – ошибка в положении даже одной аминокислоты – уже ЧП, как правило, с тяжелыми и нередко летальными последствиями для клетки. Поэтому белки могут быть получены в истинно индивидуальном состоянии (в том смысле, в котором это понятие применяют для низкомолекулярных веществ). Биосинтез полисахаридов протекает по совершенно иной схеме: здесь нет матрицирования, структура и размер молекул управляются иными механизмами. Хотя в большинстве случаев мы мало знаем об этих механизмах, нам известен результат их функционирования. А он принципиально отличен от результата биосинтеза белков.
Все изученные к настоящему времени полисахариды отличаются так называемой микрогетерогенностью. Это значит, что в любом образце индивидуального (очищенного всем доступными современной науке методами) полисахарида некоторые количественные параметры, при помощи которых можно описать его структуру, варьируют в некоторых пределах. Чаще всего варьирует молекулярная масса, число разветвлений на макромолекулу, длина боковых цепей, соотношение моносахаридных остатков (если полисахарид не регулярен), степень замещения неуглеводными остатками (типа эфиров серной кислоты или метиловых эфиров). Поэтому находимые экспериментально структурные характеристики такого типа почти всегда означают результат усреднения по всем имеющимся в изученном образце молекулам полисахарида. Так, например, если анализ дает для полисахарида молекулярную массу 100000, это означает, что в образце есть молекулы с массой и 95000, и 97000, и 105000 и т.д. (разумеется эти вариации не непрерывны: «квантом» молекулярной массы в данном случае является одно моносахаридное звено). Точно так же, если установлено, что в среднем полисахарид имеет 100 разветвлений, в нем можно обнаружить молекулы и с 90, и с 91, и с 92 и т.д. разветвлениями. Истинная картина распределения этих параметров почти никогда не бывает известна (из-за исключительно больших экспериментальных трудностей). Совокупность имеющихся данных, однако, создает впечатление, что структурные вариации в пределах индивидуального полисахарида подчиняются лишь закону случая. Подчеркнем: речь идет лишь о некоторых структурных параметрах, а отнюдь не обо всех.
Интересно оценить количественно, каким может быть число структурных вариантов индивидуального полисахарида.
Рассмотрим гипотетический разветвленный полисахарид, построенный из двух типов мономерных остатков (А и Б) с такой структурой:
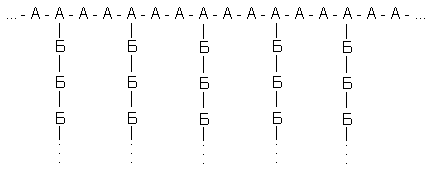
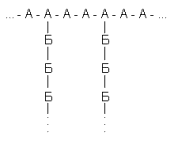
Примем условно, что у всех его молекул длина главной цепи одинакова (240 остатков), что положение разветвлений строго фиксировано (у каждого третьего), и посмотрим, что дадут вариации только одного параметра – длины боковых цепей. Наложим и на этот параметр жесткие ограничения. Пусть любая из боковых цепей может содержать либо 9, либо 10 остатков Б; причем появление цепи из 9 или 10 остатков в любом месте равновероятно. Таких цепей на молекулу, как ясно из принятых характеристик – 80. Тогда число возможных структур для полисахарида равно 2 80 » 1,2*10 24, т.е. вдвое больше, чем содержится молекул в моле вещества. Это значит, что в достаточно малом образце полисахарида может не найтись двух одинаковых молекул: все молекулы будут различаться по структуре * !
Этот вывод заметно отличается от того определения индивидуального вещества, которое мы давали вначале. И тем не менее полисахарид с такими характеристиками любой специалист по химии углеводов счел бы индивидуальным веществом, высокоочищенным, гомогенным. Заметим еще, что в нашем расчете мы не учли возможности вариации других структурных параметров, что в реальных полисахаридах число разветвлений может быть и 100, и 250 и более и что на самом деле вариации в длине полисахаридных цепей (как основной, так и боковых) могут быть гораздо шире, чем плюс-минус одно звено.
Итак, что же все-таки это такое – индивидуальный полисахарид? Может показаться, что полисахариды вообще построены «как попало» и что говорить об их структуре (в классическом значении этого слова) бессмысленно. Но это далеко не так. Отнюдь не любые структурные единицы полисахарида варьируют, размываются микрогетерогенностью. Среди них есть и консервативные, строго фиксированные. Трудность состоит, однако, в том, что комбинация консервативных и вариабельных признаков и степень вариабельности вариаблельных индивидуальны для каждого полисахарида (постольку, поскольку об этом можно судить сейчас, учитывая малую изученность микрогетерогенности, как явления). Поэтому, приступая к изучению структуры нового полисахарида, мы заранее не знаем, какие его характеристики окажутся усредненными величинами, а какие – строго детерминированными. Можно только полагать, что и консервативность, и вариабельность тех или иных характеристик строго подчинены выполнению полисахаридом его биологической функции, т.е. биологически оправданы. Вот простой пример.
Главная функция целлюлозы в растительной клетке – быть структурирующим материалом клеточной стенки. Последняя устроена весьма сложно, но приближенно может быть уподоблена армированному материалу типа стеклопластика или железобетона, в котором длинные пучки нитевидных молекул целлюлозы вплавлены в менее упорядоченный материал. Основой такой конструкции являются микрофибриллы – пачки длинных молекул. Для упаковки микрофибрилл молекула целлюлозы должна иметь вид длинного жесткого стержня, каким она в действительности и является.
Даже значительные вариации молекулярной массы целлюлозы не помешают ее молекулам образовывать микрофибриллы. Однако стоит только изменить конфигурацию одной единственной гликозидной связи на макромолекулу ( a - вместо b -), как в жестком стержне появится излом, микрофибриллы не смогут упаковаться, целлюлоза полностью утратит способность выполнять свою главную биологическую функцию. И для этого достаточно, чтобы в биосинтезе целлюлозы была допущена одна ошибка на тысячи правильно построенных гликозидных связей. Естественный отбор таких ошибок не прощает, и поэтому в этом пункте растительная клетка не ошибается никогда.
Приведенный пример характерен, так как обычно в полисахаридах именно конфигурация гликозидных связей оказывается наиболее консервативным, а молекулярная масса – наиболее вариабельным параметром структуры. Однако подчеркнем еще раз: в каждом типе полисахаридов могут быть свои консервативные и вариабельные элементы. Вопрос в том, что именно нужно для биологической функции. Но вот этого-то мы чаще всего и не знаем, и в конечном итоге именно для выяснения этого вопроса и работают исследователи структуры полисахаридов.