Биосинтез сложных липидов
А. Биосинтез жиров и фосфолипидов
Сложные липиды, такие, как нейтральные жиры (триацилглицерины), фосфо- и гликолипиды, синтезируются по основным реакционным путям. Синтез начинается (на схеме внизу) с sn-З-глицерофосфата. Обозначение «sn» относится к стереохимической номенклатуре, так как симметричный глицерин после присоединения фосфата приобретает хиральность. sn-3-Глицерофосфат образуется при восстановлении [1] промежуточного продукта гликолиза, дигидроксиацетон-3-фосфата, или в результате фосфорилирования глицерина [2]. При этерификации sn-3-глицерофосфата по C-1 длинноцепочечной жирной кислотой образуются лизофосфатиды [3], при повторной этерификации ненасыщенной жирной кислотой по С-2 — фосфатидаты [4], ключевые промежуточные продукты в биосинтезе жиров, фосфо- и гликолипидов.
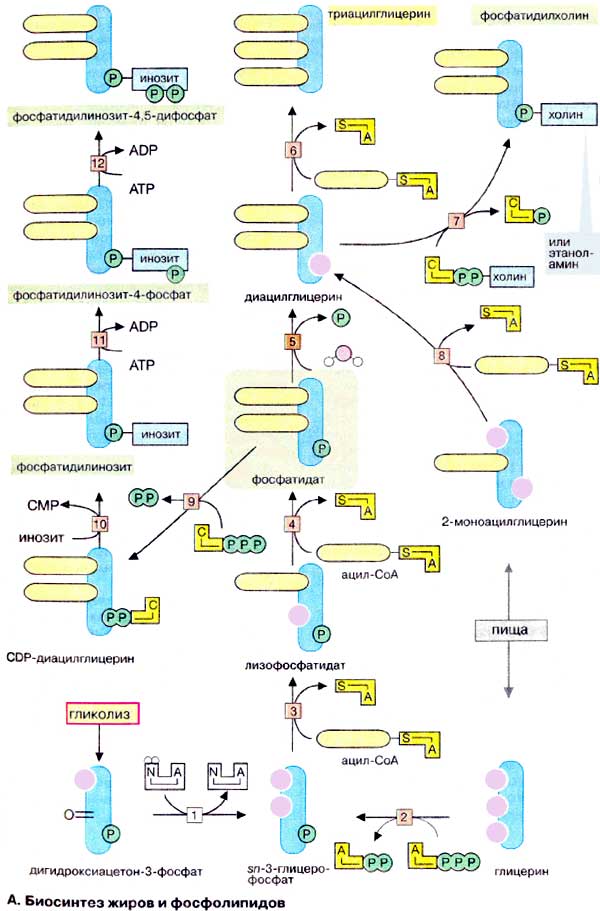
Из фосфатидовых кислот после гидролитического отщепления фосфатной группы [5] и последующего ацилирования жирной кислотой [6] образуются триацилглицерины (жиры). После завершения биосинтеза нейтральные жиры сохраняются в цитоплазме в виде жировых капель.
Из фосфатидовых кислот также через промежуточное образование диацилглицерина синтезируется фосфатидилхолин (лецитин) [7]. Фосфохолиновая группа переносится на диацилглицерин из холинцитидиндифосфата [ЦДФ-холина (CDP-холина)], представляющего собой активированную форму холина.
По аналогичной реакции диацилглицерина и ЦДФ-этаноламина образуется фосфатидилэтаноламин. Фосфатидилсерин образуется из фосфатидилэтаноламина путем обмена аминоспиртовых групп. Последующие реакции заключаются во взаимопревращении фосфолипидов: например, фосфатидилсерин декарбоксилируется с образованием фосфатидилэтаноламина; последний в результате метилирования S-аденозилметионином превращается в фосфатидилхолин.
Для биосинтеза нейтральных жиров, фосфо- и гликолипидов также используются пищевые липиды. Нейтральные жиры расщепляются в пищеварительном тракте панкреатическими липазами до жирных кислот и 2-моноацилглицеринов. Эти метаболиты всасываются слизистой кишечника и используются в качестве предшественников в синтезе липидов. Последовательное ацилирование 2-моноацилглицерина с образованием в качестве конечных продуктов нейтральных жиров катализируется двумя ацилтрансферазами [8, 6] в последовательности реакций: 2-моноацилглицерин → диацилглицерин → триацилглицерин (нейтральный жир).
Биосинтез фосфатидилинозита также начинается с фосфатидовой кислоты. При взаимодействии с ЦМФ (CMP) образуется ЦДФ-диацилглицерин [9], который далее реагирует с инозитом с образованием фосфатидилинозита [10]. Этот фосфолипид переходит в плазматическую мембрану и может быть там фосфорилирован АТФ с образованием фосфатидилинозит-4-фосфата (PlnsP) и фосфатидилинозит-4,5-дифосфата (PlnsP2). PlnsP2 является субстратом фосфолипазы С, которая расщепляет его до вторичных мессенджеров 2,3-диацилглицерина [ДАГ (DAG)] и инозит-1,4,5-трифосфата [ИФ3 (InsP3)] (см. рис. 375).
Ниже перечислены ферменты, принимающие участие в биосинтезе липидов. Большинство из них ассоциировано с мембранами гладкого эндоплазматического ретикулума, где и протекают представленные на схеме реакции.
[ 1] 3-Глицерофосфатдегидрогеназа (NAD+) 1.1.1.8[ 2] Глицеринкиназа 2.7.1.30
[ 3] 3-Глицерофосфат-О-ацилтрансфераза 2.3.1.15
[ 4] 1-Ацил-3-глицерофосфат-О-ацилтрансфераза 2.3.1.51
[ 5] Фосфатидатфосфатаза 3.1.3.4
[ 6] Диацилглицерин-0-ацилтрансфераза 2.3.1.20
[ 7] 1-Алкил-2-ацетилглицерин—холинфосфотрансфераза 2.7.8.16
[ 8] Ацилглицерин-О-пальмитоилтрансфераза 2.3.1.22
[ 9] Фосфатидат—цитидилтрансфераза 2.7.7.41
[10] CDP-диацилглицерин—инозит-3-фосфатидилтрансфераза 2.7.8.11
[11] 1-Фосфатидилинозит-киназа 2.7.1.67
[12] 1-Фосфатидилинозит-4-фосфат-киназа 2.7.1.68