Опорные функции
Между животными клетками, с одной стороны, и растительными и бактериальными – с другой, имеется несколько кардинальных различий. К их числу относятся различия в среде обитания этих клеток. Клетки животного организма погружены в специально созданную жидкую среду – кровь или лимфу. Эти жидкости в известном смысле подобны по составу древнему Океану, в котором некогда возникла жизнь (часто говорят поэтому, что животные носят в себе частицу моря). Суммарные молярные концентрации низкомолекулярных веществ во внеклеточных жидкостях животного и в цитоплазме близки. Поэтому животные клетки находятся в осмотическом равновесии со средой, а их мембраны не подвергаются механическим нагрузкам за счет неравновесной диффузии воды внутрь клетки или из нее.
Растительные и бактериальные клетки находятся в совершенно другом положении. Внешней средой для них часто оказываются весьма разбавленные водные растворы – почти чистая вода, тогда как суммарная молярность содержимого клеток составляет величину порядка нескольких десятых. Свободная диффузия воды внутрь клетки, т.е. по градиенту концентрации, развивает в ней значительное избыточное давление (до 20 атм.), которое тонкая полужидкая мембрана выдержать не может. Поэтому такие клетки окружены жестким каркасом, называемым клеточной стенкой, который принимает на себя осмотическое давление, придает клеткам определенную устойчивую форму и защищает от внешних механических воздействий. У многоклеточных растений именно прочность клеточных стенок обусловливает способность организма сохранять постоянную форму и противостоять гравитационным, ветровым, волновым и другим механическим нагрузкам.
У животных, лишенных внутреннего скелета (беспозвоночных), выработались те или иные приспособления, выполняющие опорные функции. В частности, у членистоногих, высокоорганизованного типа беспозвоночных, тело покрыто твердой внеклеточной оболочкой (кутикулой членистоногих), выполняющей функции наружного скелета: механической защиты организма и опоры для органов движения (общеизвестным примером могут служить панцири ракообразных). По наружному расположению и основной биологической роли кутикула у членистоногих может быть уподоблена клеточной стенке.
Молекулярную основу механической прочности и стенки бактериальной клетки, и кутикулы членистоногих составляют неразветвленные полисахариды, молекулы которых имеют конформацию жесткого стержня. Такая конформация характерна для полисахаридных цепей, в которых две связи элементарного звена (моносахаридного остатка) ориентированы в пространстве параллельно. Это возможно для пиранозных звеньев, соединенных 1 à 4-связями, если и гликозидный кислород, и кислород при С-4 связаны с циклом экваториально. Одна из наиболее типичных укладок таких звеньев в стержнеобразную макромолекулу, включающая антипараллельную ориентацию соседних остатков, показана ниже:
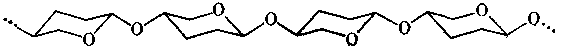
Основные типы полисахаридов, используемых живыми организмами для создания таких жестких стержневых структур, построены из 1 à 4-связанных остатков моносахаридов с β -D-глюко-кофигурацией. Это D-глюкоза в целлюлозе (растительная стенка), N-ацетил-D-глюкозамин в хитине (кутикула членистоногих) и N-ацетил-D-глюкозамин+N-ацетилмурамовая кислота в гликопептиде бактериальной стенки. При однотипности полисахаридной основы эти механические каркасы указанных групп организмов резко отличаются по принципам организации надмолекулярной структуры и по этому признаку распадаются на три класса: стенка бактериальной клетки, стенка растительной клетки, кутикула членистоногих. Мы здесь коротко разберем структуру первых двух.
Смешанный биополимер, построенный из полисахаридных цепей и коротких пептидных фрагментов (высокомолекулярный гликопептид), составляет основу структуры стенки бактериальной клетки. Его полисахаридные цепи собраны из правильно чередующихся β -1 à 4-связанных остатков N-ацетил-D-глюкозамина (1) и N-ацетил-мурамовой кислоты (2). Последняя представляет собой простой эфир N-ацетил-D-глюкозамина (по положению 3) и молочной (L- α -оксипропионовой) кислоты.
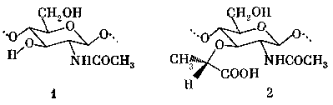
Карбоксильные группы остатка мурамовой кислоты ацилируют N-конец коротких пептидов, создающих таким образом неуглеводные разветвления полисахаридной цепи 3. В состав этих пептидов входит лишь очень ограниченный набор аминокислот, в том числе совершенно необычные для живого мира аминокислоты D-ряда (D-аланин 4 и D-глютаминовая кислота 5). Обязательными компонентами таких пептидов являются двухосновная аминокислота (D-глютаминовая) и аминокислоты, содержащие две аминогруппы: L-лизин (6) или эритро-(7) и D-трео- α , e -диаминопимелиновая кислота (8). Эта особенность позволяет пептидным цепям в свою очередь нести разветвления: g -карбоксильная группа остатка глютаминовой кислоты ацилирует N-конец пентапептида, построенного из остатков глицина, а С-конец этого пентапептида ацилирует e -аминогруппу остатка лизина или диаминопимелиновой кислоты (9).
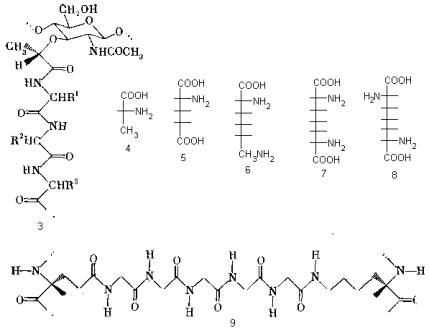
Таким образом, пентаглициновые фрагменты играют роль поперечных связок, сшивающих полисахаридные и пептидные цепи в трехмерную сетку. Общая схема образующейся структуры представлена ниже:
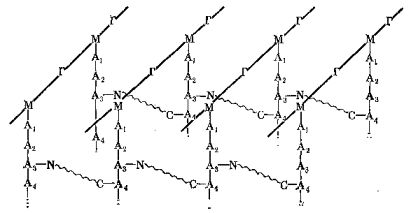
Толстые линии на ней – это полисахаридные цепи, М и Г – остатки N-ацетилмурамовой кислоты и N-ацетил-D-глюкозамина соответственно; тонкие вертикальные линии – пептидные цепи (N-конец наверху), А 1, А 2 и т.д. - остатки аминокислот, волнистые линии – пентапептидный фрагмент из остатков глицина, латинскими буквами обозначены его N- и C-концы.
Весь этот гликопептид стенки представляет собой одну гигантскую макромолекулу в форме мешка, имеющую молекулярную массу порядка десятков миллиардов дальтон. Высокая механическая прочность этой системы обусловлена сшитой структурой и жесткостью стрержнеобразных полисахаридных цепей, составляющих каркас всей конструкции. Помимо своей основной механической функции, гликопептид клеточной стенки принимает участие и в ряде других важных биологических феноменов. Упомянем кратко некоторые из них.
Биосинтез гликопептида стенки проходит через несколько этапов, включающих образование полисахаридных цепей, наращивание на них пептидных разветвлений и в заключение – сшивание этих пептидов пентаглициновыми мостиками. Ряд антибиотиков блокирует определенные стадии этого процесса, что в итоге приводит к нарушению биосинтеза стенки и, следовательно, к появлению нежизнеспособных бактериальных клеток после деления. Так, бацетрацин и ванкомицин ингибируют биосинтез полисахаридных цепей гликопептида, а пенициллин угнетает заключительный этап – образование пентаглициновых сшивок. Гликопептид рассматриваемого типа – общая основа клеточной стенки самых разнообразных бактерий; в то же время подобные структуры отсутствуют в клетках животных организмов. Отсюда становятся понятными причины широты антибактериального спектра таких антибиотиков, с одной стороны, и их исключительно низкая токсичность для животных, с другой.
Полисахаридные цепи гликопротеида стенки химически весьма устойчивы. Тем не менее, их гидролиз легко протекает под действием специфического фермента – лизоцима, весьма распространенного в живых организмах. Обработка многих бактерий лизоцимом приводит к разрушению стенки и в обычных условиях к гибели бактериальной клетки (из-за способности лизировать, т.е. растворять бактериальные клетки, фермент и получил свое название). Ряд слизистых выделений животных организмов, таких, как слезы или слюна, содержит лизоцим, что обусловливает их защитный эффект против вторжения инфекции.
Стенка растительной клетки, в отличие от гликопротеинового каркаса бактериальной, построена на иных принципах
структурной организации. Растительная стенка – надмолекулярная структура, отдельные компоненты которой не связаны ковалентными связями, а держатся вместе за счет межмолекулярных взаимодействий, в которых доминируют водородные связи. По композиции эта структура может быть уподоблена армированным материалам типа железобетона или стеклопластика, а по сложной многослойной конструкции более всего напоминает автомобильную покрышку (если искать аналогии среди знакомых макроскопических объектов). Механическую основу стенки (ее корд) составляют микрофибриллы целлюлозы – пучки тесно связанных линейных макромолекул, а цементирующим материалом служат полисахариды других классов – гемицеллюлозы и пектиновые вещества, а также аморфный ароматический полимер – лигнин.
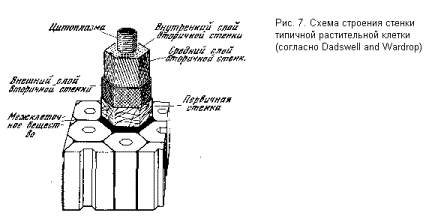
Типичная стенка растительной клетки (рис. 7) состоит из нескольких слоев. В ней различают первичную стенку, составляющую наружный слой клетки, и вторичную стенку, состоящую из внутреннего, среднего и внешнего слоев. В первичной стенке микрофибриллы не имеют определенной ориентации и переплетены в беспорядочную сеть. Во внешнем слое вторичной стенки они образуют два семейства параллельных линий, пересекающихся почти под прямым углом и образующих, таким образом, правильную («декартову») сетку. В среднем слое вторичной стенки микрофибриллы параллельны друг другу и почти параллельны оси цилиндра (фигуры, в грубом приближении
описывающей геометрию типичной растительной клетки). Наконец, во внутреннем слое вторичной стенки микрофибриллы также параллельны, но ориентированы под углом к оси. Такая сложная композиция обеспечивает исключительно высокие прочностные характеристики стенки растительной клетки (а следовательно, и растительных материалов, применяемых человеком).
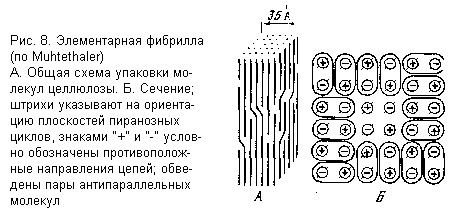
Общая схема строения микрофибрилл в настоящее время выяснена довольно полно (главным образом с помощью электронной микроскопии и рентгеноструктурного анализа), хотя целый ряд подробностей еще продолжает дискутироваться. Микрофибриллы представляют собой агрегаты из нескольких так называемых элементарных фибрилл, в которых молекулы целлюлозы вытянуты продольно, а в поперечном направлении плотно упакованы в высоко упорядоченную кристаллическую структуру. Элементарная фибрилла (рис. 8) представлена стержнем с почти квадратным сечением (угол при вершине 86,5 ° ) и стороной 35 Ǻ. На сечение приходится 36 цепей целлюлозы. В поперечном сечении элементарной фибриллы молекулы целлюлозы упакованы в правильную решетку и соединены между собой водородными связями. Соседние молекулы ориентированы антипараллельно, т.е. направление гликозидных связей у них противоположно. Примыкающие одна к другой антипараллельные цепи целлюлозы организованы в пары, между которыми образуются особенно прочные водородные связи. У концов отдельных молекул возникают дислокации, в которых соседние молекулы претерпевают небольшой изгиб, после чего их продолжения вновь встраиваются в кристаллическую решетку, обеспечивая тем самым целостность и общую упорядоченность элементарной фибриллы.
Внутри кристаллических областей элементарной фибриллы молекулы целлюлозы имеют строго упорядоченную конформацию жесткого стержня, в котором соседние глюкопиранозные остатки повернуты один относительно другого на 180 ° . Такая конформация закреплена внутримолекулярными водородными связями, соединяющими каждую пару соседних остатков. Гидроксильная группа при С-3 выступает в роли донора протона, а кислород соседнего слева остатка – в роли акцептора. Фрагмент такой структуры представлен ниже (10):
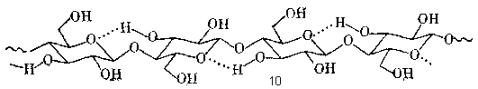
Помимо строго упорядоченных кристаллических участков, структура которых была только что рассмотрена, в нативных микрофибриллах имеются также и аморфные (или, по крайней мере, менее упорядоченные) участки, где целлюлозные цепи упакованы более рыхло. Вопрос об относительном содержании аморфных и кристаллических участков и об их взаимном расположении в элементарных фибриллах и микрофибриллах до сих пор остается дискуссионным из-за отсутствия бесспорных методов для прямого экспериментального определения этих характеристик. Однако факт существования в нативных целлюлозах двух типов надмолекулярных структур не вызывает сомнения. Такие участки резко различаются по реакционной способности, что непосредственно проявляется в химических экспериментах.
Действительно, в кристаллических участках молекулы целлюлозы упакованы плотно и с большой энергией межмолекулярной связи. Поэтому молекулы, находящиеся внутри такого участка, недоступны для реагентов и могут вступать в какие бы то ни было реакции лишь после разрушения или удаления молекул, лежащих на поверхности и вступающих в прямой контакт с реакционной средой. В этом смысле кристаллические участки ведут себя в гетерогенных реакциях как классическое твердое тело. В аморфных участках отрезки молекул упакованы менее плотно и более подвижны относительно друг друга. Внутрь этих участков может проникать и растворитель, и реагент; может происходить набухание. В результате образуются локальные гелевые структуры с большим содержанием жидкой фазы, соприкасающейся со всеми или с большинством молекул целлюлозы. Поэтому реакции полисахарида в таких участках по кинетическим характеристикам приближаются к реакциям в растворе, т.е. могут протекать значительно быстрее, чем истинно гетерогенные реакции в кристаллических участках. Так, например, при осторожном кислотном гидролизе нативной целлюлозы разрыв гликозидных связей происходит в первую очередь и почти исключительно в аморфных участках. Таким путем можно, в частности, получить препаративно высоко кристаллическую целлюлозу, состоящую почти исключительно из кристаллических «обрезков» и непосредственно измерить среднюю длину кристаллических участков. По разным определениям (и в зависимости от источника целлюлозы) длина кристаллических участков, измеренная подобным образом, колеблется о 500 до 3300 Ǻ .
Помимо чисто научного интереса, который естественно вызывает структура такого уникального образования, как стенка растительной клетки, вопрос этот имеет крупное практическое значение. Знание тонкой структуры и подробностей формирования микрофибрилл и клеточной стенки в целом составляет солидную часть научного фондамента целлюлозной промышленности и производства натурального и искусственного волокна на основе целлюлозы. Характерным примером может служить непосредственная связь гелеобразующих свойств таких синтетических производных целлюлозы, как карбоксиметилцеллюлозы и частично метилированные целлюлозы, с распределением аморфных и кристаллических участков в исходном целлюлозном материале.