Постсинтетическая модификация белков
На V, последней, стадии синтеза белка происходят формирование третичной структуры и процессинг молекулы полипептида. Синтезированная на рибосоме в строгом соответствии с генетической программой линейная одномерная полипептидная молекула уже содержит определенную информацию. Такая молекула называется конформационной, т.е. она претерпевает не хаотичные структурные изменения, а подвергается превращению (процессингу) в строго определенное трехмерное тело, которое само наделено информацией, но уже функциональной. Указанное положение справедливо для молекул белков, выполняющих в основном структурные функции, но не для биологически неактивных молекул предшественников белков, функциональная активность которых проявляется позже в результате разнообразных превращений, объединенных понятием «постсинтетическая, или посттрансляционная, модификация». Подобные модификации структуры полипептида начинаются или сразу после трансляции, или еще до окончания формирования третичной структуры белковой молекулы.
Помимо указанного процесса протеолитического удаления сигнального пептида, во многих белках отщепляется начальный N-концевой метионин. Оказалось, что в прокариотических клетках имеются особые ферменты, модифицирующие N-концевые остатки, в частности деформилаза, катализирующая отщепление формильной группы от N-концевого метионина, а также аминопептидазы, катализирующие отщепление не только N-кон-цевого формилметионина (или метионина у эукариот), но, возможно, и других остатков аминокислот с N-конца пептида. Аналогичному так называемому ограниченному постсинтетическому протеолизу подвергаются некоторые пробелки, или проферменты (например, трипсиноген, химо-трипсиноген и др.), и предшественники гормонов (например, препроинсу-лин, пре-β-липотропин и др.). В ряде случаев наблюдается и С-концевая модификация синтезированного белка.
Как известно, участок ДНК, несущий информацию о синтезе индивидуального белка, называется геном, а участок, контролирующий синтез единственной полипептидной цепи и ответственный за него,– цистроном. Следовательно, если белок состоит из нескольких (более одного) полипептидов, то естественно предположить, что в синтезе такого белка должны участвовать несколько (более одного) цистронов. Это не всегда соответствует действительности, особенно если полипептидные цепи идентичны (например, α2- и β2-цепи гемоглобина). Если, например, пептидные цепи какой-либо одной белковой молекулы являются неидентичными, то это не всегда означает, что они синтезируются как результат действия разных цистронов. Подобный белок может синтезироваться в виде единственной полипептидной цепи с последующими протеолитическими разрывами в одном или нескольких местах и отщеплением неактивных участков. Типичным примером подобной модификации является гормон инсулин, синтезирующийся в виде единого полипептида препроинсулина, который после ферментативного гидролиза превращается сначала в неактивный предшественник проинсулин, а затем в активный гормон инсулин, содержащий две разных размеров и последовательности полипептидные цепи (см. рис. 1.14).
Следует подчеркнуть, однако, что значительно больший удельный вес имеет посттрансляционная химическая модификация белков, затрагивающая радикалы отдельных аминокислот. Одной из таких существенных модификаций является ковалентное присоединение простетической группы к молекуле белка. Например, только после присоединения пи-ридоксальфосфата к ε-аминогруппе остатка лизина белковой части – апо-ферменту – образуется биологически активная трехмерная конфигурация аминотрансфераз, катализирующих реакции трансаминирования аминокислот. Некоторые белки подвергаются гликозилированию, присоединяя олигосахаридные остатки (образование гликопротеинов), и обеспечивают тем самым доставку белков к клеткам-мишеням. Широко представлены химические модификации белков в результате реакции гидроксили-рования остатков пролина, лизина (при формировании молекул коллагена), реакции метилирования (остатки лизина, глутамата), ацети-лирования ряда N-концевых аминокислот, реакции карбоксилиро-вания остатков глутамата и аспартата ряда белков (добавление экстракарбоксильной группы). В частности, протромбин (белок свертывающей системы крови) содержит ряд γ-карбоксиглутаматных остатков на N-конце, в образовании которых активное участие принимает витамин К, содержащий фермент. Предполагают, что γ-карбоксиглутаматные остатки принимают участие в связывании ионов Са2+, необходимых для инициации свертывания крови.
Одной из широко распространенных химических постсинтетических модификаций является фосфорилирование остатков серина и треонина, например, в молекуле гистоновых и негистоновых белков, а также казеина молока. Фосфорилирование-дефосфорилирование ОН-группы серина абсолютно необходимо для множества ферментов, например для активности гликоген-фосфорилазы и гликоген-синтазы. Фосфорилирование некоторых остатков тирозина в молекуле белка в настоящее время рассматривается как один из возможных и специфических этапов формирования онкобелков при малигнизации нормальных клеток. Хорошо известны также реакции окисления двух остатков цистеина и образование внутри- и межцепочечных дисульфидных связей при формировании третичной структуры (фолдинг). Этим обеспечивается не только защита от внешних денатурирующих агентов, но и образование нативной конформации и проявление биологической активности.
Менее известны реакции фарнезилирования остатков цистеина ряда белков: белка G (см. главу 8), группы белков ядерного матрикса, а также белков-онкогенов ras и протоонкогенов; источником изопрениль-ных групп является фарнезил-пирофосфат (промежуточный продукт при синтезе холестерина). Получены доказательства, что блокирование реакции фарнезилирования, вызванное специфическими препаратами (ингибиторами), приводит к потере канцерогенной активности онкогена ras. Эти результаты могут служить основой для разработки эффективных средств борьбы с опухолевыми заболеваниями человека, основанными на инги-бировании посттрансляционной модификации белков вообще или онко-белков в частности.
Следует отметить, что, хотя биосинтез белка, представляющий сложный многоступенчатый процесс, подробно описан во многих обзорах и монографиях, наши знания о структурно-функциональных взаимоотношениях многих его этапов все еще недостаточны. Действительно, выделены и охарактеризованы рибосомы (более полно у Е. coli), состоящие из множества индивидуальных белков и 3 типов молекул РНК; более того, выяснена аминокислотная последовательность всех 55 белковых молекул, первичная и вторичная структура 3 типов РНК, интенсивно изучается трехмерная структура отдельных белков рибосом прокариот. Тем не менее многие существенные детали механизма белкового синтеза неясны. Например, недостаточно известно, какие участки или составные части рибосом ответственны за инициацию, элонгацию и терминацию белкового синтеза; каков молекулярный механизм процессов транслокации, пептидилтранс-феразной реакции; каковы тонкие взаимодействия рибосом с белковыми факторами, мРНК, тРНК и антибиотиками. Потребуется еще немало усилий для определения полной молекулярной архитектуры рибосом и отдельных ее субчастиц, а также для выяснения и получения точных данных об их третичной структуре, форме и размерах, достаточных для раскрытия на молекулярном уровне функций рибосомы в сложном процессе синтеза белка.
Внерибосомный механизм синтеза пептидов. Накопленные данные, действительно, свидетельствуют о том, что матричный механизм синтеза лежит в основе биосинтеза почти всех белков живых организмов. Тем не менее синтез ряда низкомолекулярных (коротких) пептидов в биологических системах может осуществляться не только без участия нуклеиновых кислот, в частности без матричной мРНК, но даже в отсутствие рибосом. Еще на X Международном биохимическом конгрессе в Гамбурге в 1976 г. Ф. Липман (США) и К. Курахаси (Япония) представили экспериментальные доказательства синтеза двух природных циклических пептидных антибиотиков – грамицидина S и тироцидина как в цельных экстрактах, полученных из Bacillus brevis, так и в изолированных из экстрактов белковых фракциях. В частности, выделенные из экстрактов В. brevis и очищенные два белковых препарата обеспечивали точность сборки циклического полипептида – грамицидина S, состоящего из 10 аминокислотных остатков, расположенных в строгой последовательности. Очищенные белковые фракции (с мол. массами 100000 и 180000) требовали присутствия только свободных аминокислот, АТФ и ионов Mg2+для синтеза этого циклического декапептида (О-фенилаланилпролилвалилорнитиллейцин)2:
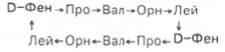
Показано, что именно легкая белковая фракция (мол. масса 100000) обеспечивает рацемизирование и включение D-фенилаланина в первую полипептидную цепь, а тяжелая фракция (мол. масса 180000) – включение 4 остальных L-аминокислот; оба фермента принимают участие также и в образовании пептидных связей. Аналогично синтезируется такой же пентапептид на расположенном рядом мультиферментном комплексе, затем оба пентапептида соединяются по типу «голова» к «хвосту» с замыканием цепи и образованием циклического декапептида. В механизме синтеза предполагается предварительное образование аминоациладенилатов (при участии этих же ферментов), из которых остатки аминокислот затем переносятся на SH-группы обоих ферментов. При этом образуются активированные промежуточные тиоэфиры, подобные тиоэфирам при синтезе высших жирных кислот (см. главу 11). В структуре первого (легкого) фермента открыт ковалентно связанный остаток фосфопантотеина, поэтому предполагают участие его тиоловой группы в переносе растущей пептидной цепи с одного участка фермента на другой. Аналогичный механизм синтеза доказан также для антибиотика тироцидина (декапептид) и для 13-членного циклического пептида – антибиотика микобациллина.
Таким образом, природа (условно в лице бактериальной клетки), очевидно, не утратила полностью существовавшего до матричного, рибо-сомного, пути атавистического механизма синтеза белковых тел и пользуется для этого весьма примитивными, но достаточно эффективными приемами.